Изображение предметов на сетчатке глаза. Строение человеческого глаза
Вспомогательный аппарат зрительной системы и его функции
Зрительная сенсорная система снабжена сложным вспомогательным аппаратом, который включает глазное яблоко и три пары мышц, обеспечивающих его движения. Элементы глазного яблока осуществляют первичное преобразование светового сигнала, попадающего на сетчатку:
оптическая система глаза фокусирует изображения на сетчатке;
зрачок регулирует количество падающего на сетчатку света;
мышцы глазного яблока обеспечивают его непрерывные перемещения.
Формирование изображения на сетчатке
Естественный свет, отраженный от поверхности предметов, является рассеянным, т.е. световые лучи от каждой точки объекта исходят в разных направлениях. Поэтому в отсутствие оптической системы глаза лучи от одной точки объекта (а ) попадали бы в разные участки сетчатки (а1, а2, а3 ). Такой глаз смог бы различать общий уровень освещенности, но не контуры предметов (рис.1 А).
Для того, чтобы увидеть объекты окружающего мира, необходимо, чтобы световые лучи от каждой точки объекта попадали только в одну точку сетчатки, т.е. необходимо сфокусировать изображение. Этого можно добиться, поместив перед сетчаткой сферическую преломляющую поверхность. Световые лучи, исходящие из одной точки (а ), после преломления на такой поверхности будут собираться в одной точке а1 (фокусе). Таким образом, на сетчатке возникнет четкое перевернутое изображение (рис. 1 Б).
Преломление света осуществляется на границе раздела двух сред, имеющих разные коэффициенты преломления. В глазном яблоке находится 2 сферические линзы: роговица и хрусталик. Соответственно, имеется 4 преломляющие поверхности: воздух/роговица, роговица/водянистая влага передней камеры глаза, водянистая влага/хрусталик, хрусталик/стекловидное тело.
Аккомодация
Аккомодация – настройка преломляющей силы оптического аппарата глаза на определенное расстояние до рассматриваемого объекта. Согласно законам рефракции, если луч света падает на преломляющую поверхность, то он отклоняется на угол, зависящий от угла его падения. При приближении объекта, угол падения исходящих от него лучей будет изменяться, поэтому преломленные лучи соберутся в другой точке, которая будет находиться позади сетчатки, что приведет к «размытию» изображения (рис 2 Б). Для того, чтобы его вновь сфокусировать, необходимо увеличить преломляющую силу оптического аппарата глаза (рис 2 В). Это достигается увеличением кривизны хрусталик, которое происходит при повышении тонуса цилиарной мышцы.
Регуляция освещенности сетчатки
Количество света, падающего на сетчатку, пропорционально площади зрачка. Диаметр зрачка у взрослого человека изменяется от 1.5 до 8 мм, что обеспечивает изменение интенсивности падающего на сетчатку света примерно в 30 раз. Зрачковые реакции обеспечиваются двумя системами гладких мышц радужной оболочки: при сокращении кольцевых мышц зрачок сужается, при сокращении радиальных мышц – расширяется.
При уменьшении просвета зрачка резкость изображения увеличивается. Это происходит потому, что сужение зрачка препятствует попаданию света на периферические области хрусталика и тем самым устраняет искажение изображения, возникающие за счет сферической аберрации.
Движения глаз
Глаз человека приводится в движение шестью глазными мышцами, которые иннервируются тремя черепномозговыми нервами – глазодвигательным, блоковым и отводящим. Эти мышцы обеспечивают два типа перемещений глазного яблока - быстрые скачкообразные (саккады) и плавные следящие движения.
Скачкообразные движения глаз (саккады ) возникают при рассматривании неподвижных предметов (рис. 3). Быстрые повороты глазного яблока (10 - 80 мс) чередуются с периодами неподвижной фиксации взгляда в одной точке (200 - 600мс). Угол поворота глазного яблока в течение одной саккады колеблется от нескольких угловых минут до 10° , а при переводе взгляда с одного объекта на другой может достигать 90° . При больших углах смещения саккады сопровождаются поворотом головы; смещение глазного яблока обычно опережает движение головы.
Плавные движения глаз сопровождают перемещающиеся в поле зрения объекты. Угловая скорость таких движений соответствует угловой скорости объекта. Если последняя превышает 80° /с, то слежение становится комбинированным: плавные движения дополняются саккадами и поворотами головы.
Нистагм - периодическое чередование плавных и скачкообразных движений. Когда едущий в поезде человек смотрит в окно, его глаза плавно сопровождают перемещающийся за окном пейзаж, а затем взгляд скачкообразно перемещается на новую точку фиксации.
Преобразование светового сигнала в фоторецепторах
Типы фоторецепторов сетчатки и их свойства
В сетчатке имеется два типа фоторецепторов (палочки и колбочки), которые различаются строением и физиологическими свойствами.
Таблица 1. Физиологические свойства палочек и колбочек
Палочки |
Колбочки |
|
| Светочувствительный пигмент | Родопсин |
Йодопсин |
| Максимум поглощения пигмента | Имеет два максимума – один в видимой части спектра (500 нм), другой – в ультрафиолетовой (350 нм) |
Существуют 3 вида йодопсинов, которые имеют различные максимумы поглощения: 440 нм (синий), 520 нм (зеленый) и 580 нм (красный) |
| Классы клеток | Каждая колбочка содержит только один пигмент. Соответственно, существуют 3 класса колбочек, чувствительных свету с разной длиной волны | |
| Распределение по сетчатке | В центральной части сетчатки плотность палочк составляет около 150 000 на мм2 , по направлению к периферии она снижается до 50 000 на мм2. В центральной ямке и слепом пятне палочки отсутствуют. |
Плотность колбочек в центральной ямке достигает 150 000 на мм 2 , в слепом пятне они отсутствуют, а на всей остальной поверхности сетчатки плотность колбочек не превышает 10 000 на мм 2 . |
| Чувствительность к свету | У палочек примерно в 500 раз выше, чем у колбочек |
|
| Функция | Обеспечивают черно-белое (скототопическое зрение) |
Обеспечивают цветное (фототопическое зрение) |
Теория двойственности зрения
Наличие двух фоторецепторных систем (колбочки и палочки), различающихся по световой чувствительности, обеспечивает подстройку к изменчивому уровню внешнего освещения. В условиях недостаточной освещенности восприятие света обеспечивается палочками, цвета при этом неразличимы (скототопическое зрени е ). При ярком освещении зрение обеспечивается главным образом колбочками, что позволяет хорошо различать цвета (фототопическое зрение ).
Механизм преобразования светового сигнала в фоторецепторе
В фоторецепторах сетчатки осуществляется преобразование энергии электромагнитного излучения (света) в энергию колебаний мембранного потенциала клетки. Процесс преобразования протекает в несколько этапов (рис. 4).
На 1-м этапе фотон видимого света, попадая в молекулу светочувствительного пигмента, поглощается p -электронами сопряженных двойных связей 11-цис -ретиналя, при этом ретиналь переходит в транс -форму. Стереомеризация 11-цис -ретиналя вызывает конформационные изменения белковой части молекулы родопсина.
На 2-м этапе происходит активация белка трансдуцина, который в неактивном состоянии содержит прочно связанный GDP . После взаимодействия с фотоактивированным родопсином трансдуцин обменивает молекулу GDP на GTP .
На 3-м этапе GTP-содержащий трансдуцин образует комплекс с неактивной cGMP-фосфодиэстеразой, что приводит к активации последней.
На 4-м этапе активированная cGMP-фосфодиэстераза осуществляет гидролиз внутриклеточного с GMP до GMP .
На 5-м этапе падение концентрации cGMP приводит к закрытию катионных каналов и гиперполяризации мембраны фоторецептора.
В ходе трансдукции сигнала по фосфодиэстеразному механизму происходит его усиление. За время фоторецепторного ответа одна единственная молекул возбужденного родопсина успевает активировать несколько сот молекул трансдуцина. Т.о. на первом этапе трансдукции сигнала происходит усиление в 100 -1000 раз. Каждая активированная молекула трансдуцина активирует лишь одну молекулу фосфодиэстеразы, зато последняя катализирует гидролиз нескольких тысяч молекул с GMP . Т.о. на этом этапе сигнала усиливается еще в 1 000 -10 000раз. Следовательно, при передаче сигнала от фотона до cGMP может происходить более чем 100 000-кратное его усиление.
Обработка информации в сетчатке
Элементы нейронной сети сетчатки и их функции
Нейронная сеть сетчатки включает 4 типа нервных клеток (рис.5):
ганглиозные клетки,
биполярные клетки,
амакриновые клетки,
горизонтальные клетки.
Ганглиозные клетки – нейроны, аксоны которых в составе зрительного нерва выходят из глаза и следуют в ЦНС. Функция ганглиозных клеток – проведение возбуждения из сетчатки в ЦНС.
Биполярные клетки соединяют рецепторные и ганглиозные клетки. От тела биполярной клетки отходят два разветвленных отростка: один отросток образует синаптические контакты с несколькими фоторецепторными клетками, другой – с несколькими ганглиозными клетками. Функция биполярных клеток – проведение возбуждения от фоторецепторов к ганглиозным клеткам.
Горизонтальные клетки соединяют расположенные рядом фоторецепторы. От тела горизонтальной клетки отходит несколько отростков, которые образуют синаптические контакты с фоторецепторами. Основная функция горизонтальных клеток – осуществление латеральных взаимодействий фоторецепторов.
Амакриновые клетки расположены подобно горизонтальным, но их образуют контакты не с фоторецепторными, а с ганглиозными клетками.
Распространение возбуждения в сетчатке
При освещении фоторецептора в нем развивается рецепторный потенциал, который представляет собой гиперполяризацию. Рецепторный потенциал, возникший в фоторецепторной клетке, передается биполярным и горизонтальным клеткам через синаптические контакты с помощью медиатора.
В биполярной клетке может развиваться как деполяризация, так и гиперполяризация (подробнее см. ниже), которая через синаптический контакт распространяется на ганглиозные клетки. Последние являются спонтанно активными, т.е. непрерывно генерируют потенциалы действия с определенной частотой. Гиперполяризация ганглиозных клеток приводит к снижению частоты нервных импульсов, деполяризация – к ее увеличению.
Электрические реакции нейронов сетчатки
Рецептивное поле биполярной клетки представляет собой совокупность фоторецепторных клеток, с которым она образует синаптические контакты. Под рецептивным полем ганглиозной клетки понимают совокупность фоторецепторных клеток, с которыми данная ганглиозная клетка соединена через биполярные клетки.
Рецептивные поля биполярных и ганглиозных клеток имеют круглую форму. В рецептивном поле можно выделить центральную и периферическую часть (рис. 6). Граница между центральной и периферической часть рецептивного поля является динамичной и может смещаться при изменении уровня освещенности.
Реакции нервных клеток сетчатки при освещении фоторецепторов центральной и периферической части их рецептивного поля, как правило, противоположны. При этом существует несколько классов ганглиозных и биполярных клеток (ON -, OFF -клетки), демонстрирующих разные электрические ответы на действие света (рис. 6).
Таблица 2. Классы ганглиозных и биполярных клеток и их электрические реакции
Классы клеток |
Реакция нервных клеток при освещении фоторецепторов, находящихся |
|
в центральной части РП |
в периферической части РП |
|
Биполярные клетки ON типа |
Деполяризация |
Гиперполяризация |
Биполярные клетки OFF типа |
Гиперполяризация |
Деполяризация |
Ганглиозные клетки ON типа |
||
Ганглиозные клетки OFF типа |
Гиперполяризация и снижение частоты ПД |
Деполяризация и увеличение частоты ПД |
Ганглиозные клетки ON - OFF типа |
Дают короткий ON -ответ на стационарный световой стимул и короткую OFF -реакцию на ослабление света. |
|
Обработка зрительной информации в ЦНС
Сенсорные пути зрительной системы
Миелиновые аксоны ганглиозных клеток сетчатки направляются в головной мозг в составе двух зрительных нервов (рис.7). Правый и левый зрительные нервы сливаются у основания черепа, образуя зрительный перекрест (хиазму). Здесь нервные волокна, идущие от медиальной половины сетчатки каждого глаза переходят на контрлатеральную сторону, а волокна от латеральных половин сетчаток продолжаются ипсилатерально.
После перекреста аксоны ганглиозных клеток в составе зрительного тракта следуют в латеральные коленчатые тела (ЛКТ), где образуют синаптические контакты с нейронами ЦНС. Аксоны нервных клеток ЛКТ в составе т.н. зрительной лучистости достигают нейронов первичной зрительной коры (поле 17 по Бродману). Далее по внутрикорковым связям возбуждение распространяется во вторичную зрительную кору (поля 18б 19) и ассоциативные зоны коры.
Сенсорные пути зрительной системы организованы по ретинотопическому принципу – возбуждение от соседних ганглиозных клеток достигает соседних точек ЛКТ и коры. Поверхность сетчатки как бы проецируется на поверхность ЛКТ и коры.
Большая часть аксонов ганглиозных клеток заканчиваются в ЛКТ, часть же волокон следует в верхние бугры двухолмия, гипоталамус, претектальную область ствола мозга, ядро зрительного тракта.
Связь между сетчаткой и верхними буграми четверохолмия служит для регуляции движений глаз.
Проекция сетчатки в гипоталамус служит для сопряжения эндогенных циркадных ритмов с суточными колебаниями уровня освещенности.
Связь между сетчаткой и претектальной областью ствола исключительно важна для регуляции просвета зрачка и аккомодации.
Нейроны ядер зрительного тракта, которые также получают синаптические входы от ганглиозных клеток, связаны с вестибулярными ядрами ствола мозга. Эта проекция позволяет оценивать положение тела в пространстве на основании зрительных сигналов, а также служит для осуществления сложных глазодвигательных реакций (нистагм).
Обработка зрительной информации в ЛКТ
Нейроны ЛКТ имеют рецептивные поля округлой формы. Электрические реакции этих клеток аналогичны таковым ганглиозных клеток.
В ЛКТ существуют нейроны, которые возбуждаются при наличии в их рецептивном поле границы свет/темнота (контрастные нейроны) или при передвижении этой границы в пределах рецептивного поля (детекторы движения).
Обработка зрительной информации в первичной зрительной коре
В зависимости от реакции на световые стимулы нейроны коры подразделяют на несколько классов.
Нейроны с простым рецептивным полем. Наиболее сильное возбуждение такого нейрона происходит при освещении его рецептивного поля световой полоской определенной ориентации. Частота нервных импульсов, генерируемых таким нейроном уменьшается при изменении ориентации световой полоски (рис. 8 А).
Нейроны со сложным рецептивным полем. Максимальная степень возбуждения нейрона достигается при передвижении светового стимула в пределах ON зоны рецептивного поля в определенном направлении. Передвижение светового стимула в другом направлении или выход светового стимула за пределы ON зоны вызывает более слабое возбуждение (рис. 8 Б).
Нейроны со сверхсложным рецептивным полем. Максимальное возбуждение такого нейрона достигается при действии светового стимула сложной конфигурации. Например, известны нейроны, наиболее сильное возбуждение которых развивается при пересечении двух границ между светлым и темным в пределах ON зоны рецептивного поля (рис. 23.8 В).
Несмотря на огромно количество экспериментальных данных о закономерностях реагирования клеток на разные зрительные стимулы, к настоящему времени нет полной теории, объясняющей механизмы обработки зрительной информации в головном мозге. Мы не можем объяснить, каким образом разнообразные электрические реакции нейронов сетчатки, ЛКТ и коры обеспечивают распознавание образов и другие феномены зрительного восприятия.
Регуляция функций вспомогательного аппарата
Регуляция аккомодации. Изменение кривизны хрусталика осуществляется при помощи цилиарной мышцы. При сокращении цилиарной мышцы кривизна передней поверхности хрусталика увеличивается и преломляющая сила возрастает. Гладкомышечные волокна цилиарной мышцы иннервируются постганглионарными нейронами, тела которых располагаются в цилиарном ганглии.
Адекватным стимулом для изменения степени кривизны хрусталика является нечеткость изображения на сетчатке, которая регистрируется нейронам первичной коры. За счет нисходящих связей коры происходит изменение степени возбуждения нейронов претектальной области, что в свою очередь вызывает активацию или торможение преганглионарных нейронов глазодвигательного ядра (ядро Эдингера–Вестфаля) и постганглионарных нейронов цилиарного ганглия.
Регуляция просвета зрачка. Сужение зрачка происходит при сокращении кольцевых гладкомышечных волокон роговицы, которые иннервируются парасимпатическими постганглионарными нейронами цилиарного ганглия. Возбуждение последних происходит при высокой интенсивности падающего на сетчатку света, которая воспринимается нейронами первичной зрительной коры.
Расширения зрачка осуществляется при сокращении радиальных мышц роговицы, которые иннервируются симпатическими нейронами ВШГ. Активность последних находится под контролем цилиоспинального центра и претектальной области. Стимулом для расширения зрачка является уменьшение уровня освещенности сетчатки.
Регуляция движений глаз. Часть волокон ганглиозных клеток следуют к нейронам верхних бугров четверохолмия (средний мозг), которые связаны с ядрами глазодвигательного, блокового и отводящего нервов, нейроны которых иннервируют поперечнополосатые мышечные волокна мышц глаза. Нервные клетки верхних бугров получат синаптические входы от вестибюлярных рецепторов, проприорецепторов мышц шеи, что позволяет организму координировать движения глаз с перемещениями тела в пространстве.
Феномены зрительного восприятия
Распознавание образов
Зрительная система обладает замечательной способность распознавать объект при самых разных вариантах его изображения. Мы можем узнавать образ (знакомое лицо, букву и т. п.), когда некоторых его частей недостает, когда он содержит лишние элементы, когда он по-разному ориентирован в пространстве, имеет разные угловые размеры, повернут к нам разными сторонами и т.п. (рис. 9). Нейрофизиологичекие механизмы этого феномена в настоящее время интенсивно изучаются.
Постоянство формы и размеров
Как правило, мы воспринимаем окружающие предметы неизменными по форме и размерам. Хотя на самом деле их форма и размеры на сетчатке не являются постоянными. Например, велосипедист в поле зрения всегда кажется одинаковым по величине независимо от расстояния до него. Колеса велосипеда воспринимаются как круглые, хотя на самом деле их изображения на сетчатке могут быть узкими эллипсами. Это явление демонстрирует роль опыта в видении окружающего мира. Нейрофизиологические механизмы этого феномена в настоящее время неизвестны.
Восприятие глубины пространства
Изображение окружающего мира на сетчатке является плоским. Однако, мы видим мир объемным. Существует несколько механизмов, которые обеспечивают построение 3-мерного пространства на основании плоских изображений, сформированных на сетчатке.
Поскольку глаза расположены на некотором расстоянии друг от друга, то изображения, формирующиеся на сетчатке левого и правого глаза, несколько различаются друг от друга. Чем ближе расположен объект по отношению к наблюдателю, тем больше будут различаться эти изображения.
Перекрывание изображений также помогает оценить их взаимное расположение в пространстве. Изображение близкого предмета может перекрывать изображение удаленного, но не наоборот.
При смещении головы наблюдателя изображения наблюдаемых объектов на сетчатке также будут смещаться (явление параллакса). При одном и том же смещении головы изображения близких объектов будут смещаться сильнее, чем изображения удаленных
Восприятие неподвижности пространства
Если, закрыв один глаз, нажать пальцем на второе глазное яблоко, то мы увидим, что мир вокруг нас смещается в сторону. В обычных условиях окружающий мир неподвижен, хотя изображение на сетчатке постоянно «прыгает» за счет перемещения глазных яблок, поворотов головы, изменения положения тела в пространстве. Восприятие неподвижности окружающего пространства обеспечивается тем, что при обработке зрительных образов учитывается информация о движении глаз, движениях головы и положении тела в пространстве. Зрительная сенсорная система умеет «вычитать» собственные движения глаз и тела из перемещения изображения на сетчатке.
Теории цветового зрения
Трехкомпонентная теория
Основывается на принципе трихроматического аддитивного смешения. Согласно этой теории, три типа колбочек (чувствительны к красному, зеленому и синему цвету) работают как независимые рецепторные системы. Сравнивая интенсивность сигналов от трех типов колбочек, зрительная сенсорная система производит «виртуальное аддитивное смещение» и вычисляет истинный цвет. Авторы теории - Юнг, Максвелл, Гельмгольц.
Теория оппонентных цветов
Предполагает, что любой цвет можно однозначно описать, указав его положение на двух шкалах - «синий-желтый», «красный-зеленый». Цвета, лежащие на полюсах этих шкал, называют оппонентными. Эта теория подтверждается тем, что в сетчатке, ЛКТ и коре существуют нейроны, которые активируются, если их рецептивное поле освещают красным светом и тормозятся, если свет зеленый. Другие нейроны возбуждаются при действии желтого цвета и тормозятся при действии синего. Предполагается, что сравнивая степень возбуждения нейронов «красно-зеленой» и «желто-синей» системы, зрительная сенсорная система может вычислить цветовые характеристики света. Авторы теории - Мах, Геринг.
Таким образом, существуют экспериментальные доказательства обеих теорий цветового зрения. В настоящее время считается. Что трехкомпонентная теория адекватно описывает механизмы цветовосприятия на уроне фоторецепторов сетчатки, а теория оппеонентных цветов – механизмы цветовосприятия на уровне нейронных сетей.
Строение глаза очень сложно. Он относится к органам чувств и отвечает за восприятие света. Фоторецепторы могут воспринимать лучи света только в определенном диапазоне длины волн. В основном раздражающее влияние на глаз оказывает свет с длиной волны 400-800 нм. После этого происходит формирование афферентных импульсов, которые поступают далее в центры головного мозга. Так формируются зрительные образы. Глаз выполняет разные функции, например, он может определить форму, величину предметов, расстояние от глаза до объекта, направление движения, освещенность, окрашенность и ряд других параметров.
Преломляющие среды
В строении глазного яблока выделяют две системы. К первой относят оптические среды, которые обладают светопреломляющей способностью. Вторая система включает рецепторный аппарат сетчатки.
Светопреломляющие среды глазного яблока объединяют роговицу, жидкое содержимое передней камеры глаза, хрусталик и стекловидное тело. В зависимости от типа среды, различается коэффициент преломления. В частности, у роговицы этот показатель составляет 1,37, у стеловидного тела и жидкости передней камеры - 1,33, у хрусталика - 1,38, а у его плотного ядра - 1,4. Основным условием нормального зрения является прозрачность светопреломляющих сред.
Фокусное расстояние определяет степень преломления оптической системы, выражающейся в диоприях. Связь в данном случае обратно пропорциональная. Диоптрия подразумевает под собой силу линзы, фокусное расстояние которой составляет 1 метр. Если измерять оптическую силу в диоптриях, то для прозрачных сред глаза она составит 43 для роговицы, а для хрусталика будет изменяться в зависимости от удаленности предмета. Если пациент смотрит вдаль, то она составит 19 (а для всей оптической системы -58), а при максимальном приближении предмета - 33 (для всей оптической системы - 70).
Статическая и динамическая рефракция глаза
Рефракция - это оптическая установка глазного яблока при фокусировке на удаленных предметах.
Если глаз нормальный, то пучок параллельных лучей, идущих от бесконечно далекого предмета, преломляются таким образом, что фокус их совпадает с центральной ямкой сетчатки. Такое глазное яблоко называется эмметропическим. Однако, далеко не всегда человек может похвастаться такими глазами.
Например, близорукость сопровождается увеличением длины глазного яблока (превышает 22,5-23 мм) или увеличением преломляющей силы глаза за счет изменения кривизны хрусталика. При этом параллельный пучок света не попадает на зону макулы, а проецируется перед ней. В результате на плоскость сетчатки попадают уже расходящиеся лучи. В этом случае изображение получается расплывчатым. Глаз называют миопическим. Чтобы изображение стало четким, необходимо передвинуть фокус на плоскость сетчатки. Этого можно достичь в том случае, если пучок света имеет не параллельные, а расходящиеся лучи. Этим можно объяснить тот факт, что близорукий пациент хорошо видит вблизи.
Для контактной коррекции миопии применяют двояковогнутые линзы, способные отодвинуть фокус в зону макулы. Этим можно компенсировать повышенную преломляющую способность вещества хрусталика. Довольно часто миопия носит наследственный характер. При этом пик заболеваемости приходится на школьный возраст и связан с нарушением гигиенических правил. В тяжелых случаях миопия способна вызвать вторичные изменения сетчатки, которые могут сопровождаться значительным снижением зрения и даже слепотой. В связи с этим очень важно вовремя проводить профилактические и лечебные мероприятия, в том числе правильно питаться, заниматься физкультурой, соблюдать гигиенические рекомендации.
Дальнозоркость сопровождается уменьшением длины глаза или снижением коэффициента преломления оптических сред. При этом пучок параллельных лучей от далекого предмета попадает за плоскость сетчатки. В макуле же проецируется участок сходящихся лучей, то есть изображение получается размытым. Глаз называют при этом дальнозорким, то есть гиперметропическим. В отличие от нормального глаза, ближайшая точка ясного видения в этом случае отстоит на некоторое расстояние. Для коррекции гиперметропии можно использовать двояко выпуклые линзы, способные увеличить преломляющую силу глаза. Важно понимать, что истинная врожденная или приобретенная дальнозоркость отличается от пресбиопии (старческой дальнозоркости).
При астигматизме нарушена способность концентрировать лучи света в одной точке, то ест фокус представлен пятном. Связано это с тем, что кривизна хрусталика различается по разным меридианам. При большей преломляющей способности по вертикали, астигматизм принято называть прямым, при увеличении горизонтальной составляющей - обратным. Даже в случае нормального глазного яблока оно несколько астигматично, так как идеально ровной роговицы не бывает. Если рассматривать диск с концентрическими кругами, то возникает незначительное их сплющивание. Если астигматизм приводит к нарушению зрительной функции, то его корректируют с использованием цилиндрических линз, которые располагают в соответствующих меридианах.
Аккомодация глаза обеспечивает четкое изображение даже при разной удаленности предметов. Эта функция становится возможной, благодаря эластическим свойствам хрусталика, который свободно меняет кривизну, а, следовательно, и преломляющую силу. В связи с этим даже при перемещении объекта лучи, отраженные от него, фокусируются на плоскость сетчатки. Когда человек рассматривает бесконечно отдаленные предметы, ресничная мышца находится в расслабленном состоянии, циннова связка, которая крепится к передней и задней хрусталиковой капсуле, натянута. При натяжении волокон цинновой связки возникает растягивание хрусталика, то есть кривизна его уменьшается. При взгляде вдаль за счет наименьшей кривизны хрусталика, его преломляющая способность также наименьшая. По мере приближения предмета к глазу происходит сокращение ресничной мышцы. В результате циннова связка расслабляется, то есть хрусталик перестает растягиваться. В случае полного расслабления волокон цинновой связки хрусталик под действием силы тяжести опускается примерно на 0,3 мм. В связи эластическими свойствами хрусталиковая линза при отсутствии натяжения становится более выпуклой, а преломляющая сила ее увеличивается.

За сокращение волокон ресничной мышцы отвечает возбуждение парасимпатичесих волокон глазодвигательного нерва, которые реагируют на приток афферентных импульсов в зону среднего мозга.
Если аккомодация не работает, то есть человек смотрит вдаль, то передний радиус кривизны хрусталика составляет 10 мм, при максимальном сокращении ресничной мышцы передний радиус кривизны хрусталика изменяется до 5,3 мм. Изменения заднего радиуса менее значительные: с 6 мм он уменьшается до 5,5 мм.
Аккомодация начинает работать в тот момент, когда предмет приближается на расстояние примерно 65 метров. При этом ресничная мышца переходит из расслабленного состояния в напряженное. Однако при такой удаленности предметов напряжение волокон не велико. Более существенное сокращение мышцы возникает при приближении предмета до 5-10 метров. В дальнейшем степень аккомодации прогрессивно увеличивается до тех пор, пока предмет не выходит из зоны четкой видимости. Наименьшее расстояние, на котором предмет еще виден отчетливо, называется точкой ближайшего ясного видения. В норме дальняя точка ясного видения располагается бесконечно далеко. Интересно, что у птиц и млекопитающих механизм аккомодации сходен с человеческим.
С возрастом происходит снижение эластичности хрусталиковой линзы, при этом амплитуда аккомодации снижается. При этом дальняя точка ясного видения обычно остается на прежнем месте, а ближайшая постепенно отодвигается.
Важно отметить, что при занятиях на близком расстоянии примерно треть аккомодации остается в запасе, поэтому глаз не утомляется.
При старческой дальнозоркости происходит удаления ближайшей точки ясного видения из-за снижения эластичности хрусталика. При пресбиопии уменьшается преломляющая сила хрусталиковой линзы даже при наибольшем усилии аккомодации. В возрасте десяти лет ближайшая точка располагается в 7 см от глаза, в 20 лет смещается на 8,3 см, в 30 лет - до 11 см, к шестидесяти годам она уже сдвигается к 80-100 см.
Построение изображения на сетчатке
Глаз является очень сложной оптической системой. Для изучения его свойств используют упрощенную модель, которую называют редуцированным глазом. Зрительная ось этой модели совпадает с осью обычного глазного яблока и проходит сквозь центры преломляющих сред, попадая в центральную ямку.
В редуцированной модели глаза к преломляющим средам относят только вещество стекловидного тела, в котором отсутствуют главные точки, лежащие в области пересечения преломляющих плоскостей. В истинном глазном яблоке две узловые точки располагаются на расстоянии 0,3 мм друг от друга, их заменяют одной точкой. Луч, который проходит через узловую точку, обязательно должен пройти через сопряженную с ней, покинув ее в параллельном направлении. То есть в редуцированной модели две точки заменены одной, которая помещена на расстоянии в 7,5 мм от поверхности роговицы, то есть в задней трети хрусталика. От сетчатки узловая точка удалена на 15 мм. В случае построения изображения все точки сетчатки рассматриваются как светящиеся. От каждой из них через узловую точку проводится прямая линия.
Изображение, которое формируется на сетчатке уменьшенное, обратное и действительное. Чтобы определить размер на сетчатке, нужно зафиксировать длинное слово, которое напечатано мелким шрифтом. При этом определяют, какое количество букв может различить пациент при полной неподвижности глазного яблока. После этого линейкой измеряют длину букв в миллиметрах. Далее путем геометрических расчетов можно определить длину изображения на сетчатке. Этот размер дает представление о диаметре желтого пятна, которое отвечает за центральное четкое зрение.
Изображение на сетчатке получается обратным, но мы видим предметы прямыми. Связано это с ежедневной тренировкой головного мозга, в частности зрительного анализатора. Чтобы определить положение в пространстве, помимо раздражителей с сетчатки, человек использует возбуждение проприорецепторов мышечного аппарата глаза, а также показания других анализаторов.
Можно сказать, что формирование представлений о положении тела в пространстве основывается на условных рефлексах.
Передача зрительной информации
В последних научных исследованиях было установлено, что в процессе эволюционного развития количество элементов, которые передают информацию с фоторецепторов, увеличивается вместе с числом параллельных цепей афферентных нейронов. Это можно заметить на слуховом анализаторе, но в большей степени именно на зрительном анализаторе.
В зрительном нерве имеется около миллиона нервных волокон. Каждое волокно разделяется на 5-6 частей в промежуточном мозге и заканчивается синапсами в зоне наружного коленчатого тела. При этом каждое волокно на пути от коленчатого тела к большим полушариям головного мозга контактирует с 5000 нейронов, относящихся к зрительному анализатору. Каждый же нейрон зрительного анализатора получает информацию еще от 4000 нейронов. В результате происходит значительное расширение зрительных контактов по направлению к большим полушариям головного мозга.

Фоторецепторы в сетчатке могут передать информацию однократно в тот момент, когда появился новый предмет. Если изображение не изменяется, то в результате адаптации рецепторы перестают возбуждаться, с этим связано то, что информация о статических изображениях не передается в мозг. Также в сетчатке имеются рецепторы, которые передают только изображения предметов, другие же реагируют на движение, появление, исчезновение светового сигнала.
Во время бодрствования по зрительным нервам постоянно предаются афферентные сигналы от фоторецеторов. При разных условиях освещения эти импульсы могут возбуждаться или тормозиться. В зрительном нерве можно выделить три типа волокон. К первому типу относят волокна, которые реагируют только на включение света. Второй тип волокон приводит к торможению афферентных импульсов и реагирует на прекращение освещения. Если повторно включить освещение, то разряд импульсов в этом типе волокон будет тормозиться. Третий тип включает наибольшее количество волокон. Они реагируют как на включение, так и на выключение освещения.
При математическом анализе результатов электрофизиологических исследований установлено, что по пути от сетчатки к зрительному анализатору происходит укрупнение изображения.
Элементами зрительного восприятия являются линии. Первым делом зрительная система выделяет контуры предметов. Чтобы выделить контуры предметов, достаточно врожденных механизмов.
В сетчатке имеется временная и пространственная суммация всех зрительных раздражений, относящихся к рецептивным полям. Число их при нормальном освещении может достигать 800 тысяч, что примерно соответствует количеству волокон в зрительном нерве.
Для регуляции обмена веществ в рецепторах сетчатки имеется ретикулярная формация. Если раздражать ее электрическим током при помощи игольчатых электродов, то изменяется частота афферентных импульсов, которые возникают в фоторецепторах в ответ на вспышку света. Ретикулярная формация воздействует на фоторецепторы через тонкие эфферентные гамма-волокна, которые проникают в сетчатку, а также через проприоцепторный аппарат. Обычно через некоторое время после того, как началось раздражение сетчатки афферентная импульсация внезапно возрастает. Эффект этот может сохраняться длительное время даже после прекращения раздражения. Можно сказать, что возбудимость сетчатки значительно повышают адренергические симпатические нейроны, которые относятся к ретикулярной формации. Их характеризует большой латентный период и длительно последействие.
Рецептивные поля сетчатки представлены двумя типами. К первому относят элементы, которые кодируют самые простые конфигурации образа с учетом отдельных структур. Второй тип отвечает за кодирование конфигурации в целом, за счет их работы происходит укрупнение зрительных образов. Другими словами, статическое кодирование начинается еще на уровне сетчатки. После выхода из сетчатки импульсы поступают в зону наружных коленчатых тел, где и происходит основное кодирование зрительного образа с применением крупных блоков. Также в этой зоне передаются отдельные фрагменты конфигурации изображения, скорость и направление его движения.
На протяжении жизни происходит условно-рефлекторное запоминание зрительных образов, имеющих биологическое значение. В результате рецепторы сетчатки могут предавать отдельные зрительные сигналы, но о методах декодирования пока не известно.
Из центральной ямки выходит примерно 30 тысяч нервных волокон, при помощи которых происходит передача 900 тысяч бит информации за 0,1 секунду. За это же время в зрительной зоне больших полушарий может быть обработано не более 4 бит информации. То есть объем зрительной информации ограничен не сетчаткой, а декодированием в высших центрах зрения.
Рецептора
Афферентного проводящего пути
3) зоны коры, куда проецируется данный вид чувствительности -
И. Павлов назвал анализатором.
В современной научной литературе анализатор чаще называют сенсорной системой . В корковом конце анализатора происходят анализ и синтез полученной информации.
Зрительная сенсорная система
Орган зрения - глаз - состоит из глазного яблока и вспомогательного аппарата. Из глазного яблока выходит зрительный нерв, соединяющий его с головным мозгом.
Глазное яблоко имеет форму шара, более выпуклого спереди. Оно лежит в полости глазницы и состоит из внутреннего ядра и окружающих его трех оболочек: наружной, средней и внутренней (рис. 1).
Рис. 1. Горизонтальный разрез глазного яблока и механизм аккомодации (схема) [Косицкий Г. И., 1985] . В левой половине хрусталик (7) уплощен при рассматривании далекого предмета, а справа он стал более выпуклым за счет аккомодационного усилия при рассматривании близкого предмета 1 - склера; 2 - сосудистая оболочка; 3 - сетчатка; 4 - роговица; 5 - передняя камера; 6 - радужка; 7 - хрусталик; 8 - стекловидное тело; 9 - ресничная мышца, ресничные отростки и ресничная связка (циннова); 10 - центральная ямка; 11 - зрительный нерв
ГЛАЗНОЕ ЯБЛОКО

Наружная оболочка называется волокнистой, или фиброзной . Задний отдел ее представляет белочную оболочку, или склеру , которая защищает внутреннее ядро глаза и помогает сохранить его форму. Передний отдел представлен более выпуклой прозрачной роговицей , через которую в глаз проникает свет.
Средняя оболочка богата кровеносными сосудами и потому называется сосудистой. В ней выделяют три части:
переднюю – радужку
среднюю - ресничное тело
заднюю - собственно сосудистую оболочку .
Радужка имеет форму плоского кольца, цвет ее может быть голубой, зеленовато-серый или коричневый в зависимости от количества и характера пигмента. Отверстие в центре радужки - зрачок - способно суживаться и расширяться. Величину зрачка регулируют специальный глазные мышцы, расположенные в толще радужки: сфинктер (суживатель) зрачка и дилататор зрачка, расширяющий зрачок. Кзади от радужки находится ресничное тело - круговой валик, внутренний край которого имеет ресничные отростки . В нем заложена ресничная мышца, сокращение которой через специальную связку передается на хрусталик и он меняет свою кривизну. Собственно сосудистая оболочка - большая задняя часть средней оболочки глазного яблока, содержит черный пигментный слой, который поглощает свет.
Внутренняя оболочка глазного яблока называется сетчаткой, или сетчатой оболочкой. Это светочувствительная часть глаза, которая покрывает изнутри сосудистую оболочку. Она имеет сложное строение. В сетчатке находятся светочувствительные рецепторы - палочки и колбочки.
Внутреннее ядро глазного яблока составляют хрусталик, стекловидное тело и водянистая влага передней и задней камер глаза.
Хрусталик имеет форму двояковыпуклой линзы, он прозрачен и эластичен, расположен позади зрачка. Хрусталик преломляет входящие в глаз световые лучи и фокусирует их на сетчатке. В этом ему помогают роговица и внутриглазные жидкости. При помощи ресничной мышцы хрусталик меняет свою кривизну, принимая форму, необходимую то для "дальнего", то для "ближнего" видения.
Позади хрусталика находится стекловидное тело - прозрачная желеобразная масса.
Полость между роговицей и радужкой составляет переднюю камеру глаза, а между радужкой и хрусталиком - заднюю камеру. Они заполнены прозрачной жидкостью - водянистой влагой и сообщаются между собой через зрачок. Внутренние жидкости глаза находятся под давлением, которое определяют как внутриглазное давление. При повышении его могут возникнуть нарушения зрения. Повышение внутриглазного давления является признаком тяжелого заболевания глаз - глаукомы.
Вспомогательный аппарат глаза состоит из защитных приспособлений, слезного и двигательного аппарата.
К защитным образованиям относятся брови, ресницы и веки. Брови предохраняют глаз от пота, стекающего со лба. Ресницы, находящиеся на свободных краях верхнего и нижнего века, защищают глаза от пыли, снега, дождя. Основу века составляет соединительнотканная пластинка, напоминающая хрящ, снаружи она покрыта кожей, а изнутри - соединительной оболочкой - конъюнктивой . С век конъюнктива переходит на переднюю поверхность глазного яблока, за исключением роговицы. При сомкнутых веках образуется узкое пространство между конъюнктивой век и конъюнктивой глазного яблока - конъюнктивальный мешок.
Слезный аппарат представлен слезной железой и слезовыводящими путями . Слезная железа занимает ямку в верхнем углу латеральной стенки глазницы. Несколько ее протоков открывается в верхний свод конъюнктивального мешка. Слеза омывает глазное яблоко и постоянно увлажняет роговицу. Движению слезной жидкости в сторону медиального угла глаза способствуют мигательные движения век. Во внутреннем углу глаза слеза скапливается в виде слезного озера, на дне которого виден слезный сосочек. Отсюда через слезные точки (точечные отверстия на внутренних краях верхнего и нижнего век) слеза попадает сначала в слезные канальцы, а затем в слезный мешок. Последний переходит в носослезный проток, по которому слеза попадает в полость носа.
Двигательный аппарат глаза представлен шестью мышцами . Мышцы начинаются от сухожильного кольца вокруг зрительного нерва в глубине глазницы и прикрепляются к глазному яблоку. Выделяют четыре прямые мышцы глазного яблока (верхняя, нижняя, латеральная и медиальная) и две косые мышцы (верхняя и нижняя). Мышцы действуют таким образом, что оба глаза движутся совместно и направлены в одну и ту же точку. От сухожильного кольца начинается также мышца, поднимающая верхнее веко. Мышцы глаза исчерченные и сокращаются произвольно.
Физиология зрения
Светочувствительные рецепторы глаза (фоторецепторы) - колбочки и палочки, располагаются в наружном слое сетчатки. Фоторецепторы контактируют с биполярными нейронами, а те в свою очередь - с ганглиозными. Образуется цепочка клеток, которые под действием света генерируют и проводят нервный импульс. Отростки ганглиозных нейронов образуют зрительный нерв.
По выходе из глаза зрительный нерв делится на две половины. Внутренняя перекрещивается и вместе с наружной половиной зрительного нерва противоположной стороны направляется к латеральному коленчатому телу, где расположен следующий нейрон, заканчивающийся на клетках зрительной зоны коры в затылочной доле полушария. Часть волокон зрительного тракта направляется к клеткам ядер верхних холмиков пластинки крыши среднего мозга. Эти ядра, так же как и ядра латеральных коленчатых тел, представляют собой первичные (рефлекторные) зрительные центры. От ядер верхних холмиков начинается тектоспинальный путь, за счет которого осуществляются рефлекторные ориентировочные движения, связанные со зрением. Ядра верхних холмиков также имеют связи с парасимпатическим ядром глазодвигательного нерва, расположенным под дном водопровода мозга. От него начинаются волокна, входящие в состав глазодвигательного нерва, которые иннервируют сфинктер зрачка, обеспечивающий сужение зрачка при ярком свете (зрачковый рефлекс), и ресничную мышцу, осуществляющую аккомодацию глаза.
Адекватным раздражителем для глаза является свет - электромагнитные волны длиной 400 - 750 нм. Более короткие - ультрафиолетовые и более длинные - инфракрасные лучи глазом человека не воспринимаются.
Преломляющий световые лучи аппарат глаза - роговица и хрусталик, фокусирует изображение предметов на сетчатке. Луч света проходит через слой ганглиозных и биполярных клеток и достигает колбочек и палочек. В фоторецепторах различают наружный сегмент, содержащий светочувствительный зрительный пигмент (родопсин в Галочках и йодопсин в колбочках), и внутренний сегмент, в котором находятся митохондрии. Наружные сегменты погружены в черный пигментный слой, выстилающий внутреннюю поверхность глаза. Он уменьшает отражение света внутри глаза и участвует в обмене веществ рецепторов.
В сетчатке насчитывают около 7 млн. колбочек и примерно 130 млн. палочек. Более чувствительны к свету палочки, их называют аппаратом сумеречного зрения. Колбочки, чувствительность к свету которых в 500 раз меньше,- это аппарат дневного и цветового видения. Цветоощущение, мир красок доступен рыбам, амфибиям, рептилиям и птицам. Доказывается это возможностью выработать у них условные рефлексы на различные цвета. Не воспринимают цвета собаки и копытные животные. Вопреки прочно установившемуся представлению, что быки очень не любят красный цвет, в опытах удалось доказать, что они не могут отличить зеленого, синего и даже черного от красного. Из млекопитающих только обезьяны и люди способны воспринимать цвета.
Колбочки и палочки распределены в сетчатке неравномерно. На дне глаза, напротив зрачка, находится так называемое пятно, в центре его есть углубление - центральная ямка - место наилучшего видения. Сюда фокусируется изображение при рассматривании предмета.
В центральной ямке имеются только колбочки. По направлению к периферии сетчатки количество колбочек уменьшается, а число палочек возрастает. Периферия сетчатки содержит только палочки.
Недалеко от пятна сетчатки, ближе к носу, расположено слепое пятно. Это место выхода зрительного нерва. В этом участке нет фоторецепторов, и оно не принимает участия в зрении.
Построение изображения на сетчатке.
Луч света достигает сетчатки, проходя через ряд преломляющих поверхностей и сред: роговицу, водянистую влагу передней камеры, хрусталик и стекловидное тело. Лучи, исходящие из одной точки внешнего пространства, должны быть сфокусированы в одну точку на сетчатке, только тогда возможно ясное видение.
Изображение на сетчатке получается действительное, перевернутое и уменьшенное. Несмотря на то что изображение перевернуто, мы воспринимаем предметы в прямом виде. Это происходит потому, что деятельность одних органов чувств проверяется другими. Для нас "низ" там, куда направлена сила земного притяжения.

Рис. 2. Построение изображения в глазу, а, б - предмет: а", б" - его перевернутое и уменьшенное изображение на сетчатке; С - узловая точка, через которую лучи идут без преломления, аα - угол зрения
Острота зрения.
Остротой зрения называется способность глаза видеть раздельно две точки. Нормальному глазу это доступно, если величина их изображения на сетчатке равна 4 мкм, а угол зрения составляет 1 мин. При меньшем угле зрения ясного видения не получается, точки сливаются.
Остроту зрения определяют по специальным таблицам, на которых изображены 12 рядов букв. С левой стороны каждой строки написано, с какого расстояния она должна быть видна человеку с нормальным зрением. Испытуемого помещают на определенном расстоянии от таблицы и находят строку, которую он прочитывает без ошибок.
Острота зрения увеличивается при яркой освещенности и очень низка при слабом свете.
Поле зрения . Все пространство, видимое глазу при неподвижно устремленном вперед взоре, называют полем зрения.
Различают центральное (в области желтого пятна) и периферическое зрение. Наибольшая острота зрения в области центральной ямки. Здесь только колбочки, диаметр их небольшой, они тесно примыкают друг к другу. Каждая колбочка связана с одним биполярным нейроном, а тот в свою очередь - с одним ганглиозным, от которого отходит отдельное нервное волокно, передающее импульсы в головной мозг.
Периферическое зрение отличается меньшей остротой. Это объясняется тем, что на периферии сетчатки колбочки окружены палочками и каждая уже не имеет отдельного пути к мозгу. Группа колбочек заканчивается на одной биполярной клетке, а множество таких клеток посылает свои импульсы к одной ганглиозной. В зрительном нерве примерно 1 млн. волокон, а рецепторов в глазу около 140 млн.
Периферия сетчатки плохо различает детали предмета, но хорошо воспринимает их движения. Боковое зрение имеет большое значение для восприятия внешнего мира. Для водителей различного вида транспорта нарушение его недопустимо.
Поле зрения определяют при помощи особого прибора - периметра (рис. 133), состоящего из полукруга, разделенного на градусы, и подставки для подбородка.

Рис. 3. Определение поля зрения при помощи периметра Форстнера
Испытуемый, закрыв один глаз, вторым фиксирует белую точку в центре дуги периметра впереди себя. Для определения границ поля зрения по дуге периметра, начиная от ее конца, медленно продвигают белую марку и определяют тот угол, под которым она видна неподвижным глазом.
Поле зрения наибольшее кнаружи, к виску - 90°, к носу и кверху и книзу - около 70°. Можно определить границы цветового зрения и при этом убедиться в удивительных фактах: периферические части сетчатки не воспринимают цвета; цветовые поля зрения не совпадают для различных цветов, самое узкое имеет зеленый цвет.
Аккомодация. Глаз часто сравнивают с фотокамерой. В нем имеется светочувствительный экран - сетчатка, на которой с помощью роговицы и хрусталика получается четкое изображение внешнего мира. Глаз способен к ясному видению равноудаленных предметов. Эта его способность носит название аккомодации.
Преломляющая сила роговицы остается постоянной; тонкая, точная фокусировка идет за счет изменения кривизны хрусталика. Эту функцию он выполняет пассивно. Дело в том, что хрусталик находится в капсуле, или сумке, которая через ресничную связку прикреплена к ресничной мышце. Когда мышца расслаблена, связка натянута, она тянет капсулу, которая сплющивает хрусталик. При напряжении аккомодации для рассматривания близких предметов, чтения, письма ресничная мышца сокращается, связка, натягивающая капсулу, расслабляется и хрусталик в силу своей эластичности становится более круглым, а его преломляющая сила увеличивается.
С возрастом эластичность хрусталика уменьшается, он отвердевает и утрачивает способность менять свою кривизну при сокращении ресничной мышцы. Это мешает четко видеть на близком расстоянии. Старческая дальнозоркость (пресбиопия) развивается после 40 лет. Исправляют ее с помощью очков - двояковыпуклых линз, которые надевают при чтении.
Аномалия зрения. Встречающаяся у молодых аномалия чаще всего является следствием неправильного развития глаза, а именно его неправильной длины. При удлинении глазного яблока возникает близорукость (миопия), изображение фокусируется впереди сетчатки. Отдаленные предметы видны неотчетливо. Для исправления близорукости пользуются двояковогнутыми линзами. При укорочении глазного яблока наблюдается дальнозоркость (гиперметропия). Изображение фокусируется позади сетчатки. Для исправления требуются двояковыпуклые линзы (рис. 134).

Рис. 4. Рефракция при нормальном зрении (а), при близорукости (б) и дальнозоркости (г). Оптическая коррекция близорукости (в) и дальнозоркости (д) (схема) [Косицкий Г. И., 1985]
Нарушение зрения, называемое астигматизмом, возникает в случае неправильной кривизны роговицы или хрусталика. При этом изображение в глазу искажается. Для исправления нужны цилиндрические стекла, подобрать которые не всегда легко.
Адаптация глаза.
При выходе из темного помещения на яркий свет мы вначале ослеплены и даже можем испытывать боль в глазах. Очень быстро эти явления проходят, глаза привыкают к яркому освещению.
Уменьшение чувствительности рецепторов глаза к свету называется адаптацией. При этом происходит выцветание зрительного пурпура. Заканчивается световая адаптация в первые 4 - 6 мин.
При переходе из светлого помещения в темное происходит темновая адаптация, продолжающаяся более 45 мин. Чувствительность палочек при этом возрастает в 200 000 - 400 000 раз. В общих чертах это явление можно наблюдать при входе в затемненный кинозал. Для изучения хода адаптации существуют специальные приборы - адаптомеры.
Основы психофизиологии., М. ИНФРА-М, 1998, с.57-72, Глава 2 Отв.ред. Ю.И. Александров
2.1. Строение и функции оптического аппарата глаза
Глазное яблоко имеет шарообразную форму, что облегчает его повороты для наведения на рассматриваемый объект и обеспечивает хорошую фокусировку изображения на всей светочувствительной оболочке глаза - сетчатке. На пути к сетчатке лучи света проходят через несколько прозрачных сред роговицу, хрусталик и стекловидное тело. Определённая кривизна и показатель преломления роговицы и в меньшей мере хрусталика определяют преломление световых лучей внутри глаза. На сетчатке получается изображение, резко уменьшенное и перевернутое вверх ногами и справа налево (рис. 4.1 а). Преломляющую силу любой оптической системы выражают в диоптриях (D). Одна диоптрия равна преломляющей силе линзы с фокусным расстоянием 100 см. Преломляющая сила здорового глаза составляет 59D при рассматривании далеких и 70,5D при рассматривании близких предметов.
Рис. 4.1.
2.2. Аккомодация
Аккомодацией называют приспособление глаза к ясному видению объектов, расположенных на разном расстоянии (подобно фокусировке в фотографии). Для ясного видения объекта необходимо, чтобы его изображение было сфокусировано на сетчатке (рис. 4.1 б). Главную роль в аккомодации играет изменение кривизны хрусталика, т.е. его преломляющей способности. При рассматривании близких предметов хрусталик становится более выпуклым. Механизмом аккомодации является сокращение мышц, изменяющих выпуклость хрусталика.
2.3. Аномалии рефракции глаза
Две главные аномалии рефракции глаза близорукость (миопия) и дальнозоркость (гиперметропия). Эти аномалии обусловлены не недостаточностью преломляющих сред глаза, а изменением длины глазного яблока (рис. 4.1 в, г). Если продольная ось глаза слишком длинна (рис. 4.1 в), то лучи от далёкого объекта сфокусируются не на сетчатке, а перед ней, в стекловидном теле. Такой глаз называется близоруким. Чтобы ясно видеть вдаль, близорукий должен поместить перед глазами вогнутые стекла, которые отодвинут сфокусированное изображение на сетчатку (рис. 4.1 д). В отличие от этого, в дальнозорком глазу (рис. 4.1 г) продольная ось укорочена, и поэтому лучи от далёкого объекта фокусируются за сетчаткой, Этот недостаток может быть компенсирован увеличением выпуклости хрусталика. Однако при рассматривании близких объектов аккомодационные усилия дальнозорких людей недостаточны. Именно поэтому для чтения они должны надевать очки с двояковыпуклыми линзами, усиливающими преломление света (рис. 4.1 е).
2.4. Зрачок и зрачковый рефлекс
Зрачок - это отверстие в центре радужной оболочки, через которое свет проходит в глаз. Он повышает чёткость изображения на сетчатке, увеличивая глубину резкости глаза и устраняя сферическую аберрацию. Расширившийся при затемнении зрачок на свету быстро сужается ("зрачковый рефлекс"), что регулирует поток света, попадающий в глаз. Так, на ярком свету зрачок имеет диаметр 1,8 мм, при средней дневной освещённости он расширяется до 2,4 мм, а в темноте - до 7,5 мм. Это ухудшает качество изображения на сетчатке, но увеличивает абсолютную чувствительность зрения. Реакция зрачка на изменение освещённости имеет адаптивный характер, так как стабилизирует освещённость сетчатки в небольшом диапазоне. У здоровых людей зрачки обоих глаз имеют одинаковый диаметр. При освещении одного глаза зрачок другого тоже суживается; подобная реакция называется содружественной.
2.5. Структура и функции сетчатки
Сетчатка - это внутренняя светочувствительная оболочка глаза. Она имеет сложную многослойную структуру (рис. 4.2). Здесь расположены два вида фоторецепторов (палочки и колбочки) и несколько видов нервных клеток. Возбуждение фоторецепторов активирует первую нервную клетку сетчатки - биполярный нейрон. Возбуждение биполярных нейронов активирует ганглиозные клетки сетчатки, передающие свои импульсы в подкорковые зрительные центры. В процессах передачи и переработки информации в сетчатке участвуют также горизонтальные и амакриновые клетки. Все перечисленные нейроны сетчатки с их отростками образуют нервный аппарат глаза, который участвует в анализе и переработке зрительной информации. Именно поэтому сетчатку называют частью мозга, вынесенной на периферию.
2.6. Структура и функции слоёв сетчатки
Клетки пигментного эпителия образуют наружный, наиболее далекий от света, слой сетчатки. Они содержат меланосомы, придающие им чёрный цвет. Пигмент поглощает излишний свет, препятствуя его отражению и рассеиванию, что способствует чёткости изображения на сетчатке. Пигментный эпителий играет решающую роль в регенерации зрительного пурпура фоторецепторов после его обесцвечивания, в постоянном обновлении наружных сегментов зрительных клеток, в защите рецепторов от светового повреждения, а также в переносе к ним кислорода и питательных веществ.
Фоторецепторы. К слою пигментного эпителия изнутри примыкает слой зрительных рецепторов: палочек и колбочек. В каждой сетчатке человека находится 6-7 млн. колбочек и 110-125 млн. палочек. Они распределены в сетчатке неравномерно. Центральная ямка сетчатки - фовеа (fovea centralis) содержит только колбочки. По направлению к периферии сетчатки количество колбочек уменьшается, а количество палочек увеличивается, так что на дальней периферии имеются только палочки. Колбочки функционируют в условиях больших освещённостей, они обеспечивают дневное и цветовое зрение ; более светочувствительные палочки ответственны за сумеречное зрение.
Цвет воспринимается лучше всего при действии света на центральную ямку сетчатки, в которой расположены почти исключительно колбочки. Здесь же и наибольшая острота зрения. По мере удаления от центра сетчатки восприятие цвета и пространственное разрешение постепенно уменьшается. Периферия сетчатки, на которой находятся исключительно палочки, не воспринимает цвета. Зато световая чувствительность колбочкового аппарата сетчатки во много раз меньше, чем у палочкового. Поэтому в сумерках из-за резкого понижения колбочкового зрения и преобладания периферического палочкового зрения мы не различаем цвет ("ночью все кошки серы").
Зрительные пигменты. В палочках сетчатки человека содержится пигмент родопсин, или зрительный пурпур, максимум спектра поглощения которого находится в области 500 нанометров (нм). В наружных сегментах трёх типов колбочек (сине-, зелено- и красночувствительных) содержатся три типа зрительных пигментов, максимумы спектров поглощения которых находятся в синей (420 нм), зеленой (531 нм) и красной (558 нм) областях спектра. Красный колбочковый пигмент получил название йодопсин. Молекула зрительного пигмента состоит из белковой части (опсина) и хромофорной части (ретиналь, или альдегид витамина "А"). Источником ретиналя в организме служат каротиноиды; при их недостатке нарушается сумеречное зрение ("куриная слепота").
2.7. Нейроны сетчатки
Фоторецепторы сетчатки синаптически связаны с биполярными нервными клетками (см. рис. 4.2). При действии света уменьшается выделение медиатора из фоторецептора, что гиперполяризует мембрану биполярной клетки. От неё нервный сигнал передаётся на ганглиозные клетки, аксоны которых являются волокнами зрительного нерва.

Рис. 4.2. Схема строения сетчатки глаза:
1 - палочки; 2 - колбочки; 3 - горизонтальная клетка; 4 - биполярные клетки; 5 - амакриновые клетки; 6 - ганглиозные клетки; 7 - волокна зрительного нерва
На 130 млн. фоторецепторных клеток приходится только 1 млн. 250 тыс. ганглиозных клеток сетчатки. Это значит, что импульсы от многих фоторецепторов сходятся (конвергируют) через биполярные нейроны к одной ганглиозной клетке. Фоторецепторы, соединённые с одной ганглиозной клеткой, образуют её рецептивное поле [Хьюбел, 1990; Физиол. зрения, 1992]. Таким образом, каждая ганглиозная клетка суммирует возбуждение, возникающее в большом количестве фоторецепторов. Это повышает световую чувствительность сетчатки, но ухудшает её пространственное разрешение. Лишь в центре сетчатки (в районе центральной ямки) каждая колбочка соединена с одной биполярной клеткой, а та, в свою очередь, соединена с одной ганглиозной клеткой. Это обеспечивает высокое пространственное разрешение центра сетчатки, но резко уменьшает его световую чувствительность.
Взаимодействие соседних нейронов сетчатки обеспечивается горизонтальными и амакриновыми клетками, через отростки которых распространяются сигналы, меняющие синаптическую передачу между фоторецепторами и биполярами (горизонтальные клетки) и между биполярами и ганглиозными клетками (амакрины). Амакриновые клетки осуществляют боковое торможение между соседними ганглиозными клетками. В сетчатку приходят и центробежные, или эфферентные, нервные волокна, приносящие к ней сигналы из мозга. Эти импульсы регулируют проведение возбуждения между биполярными и ганглиозными клетками сетчатки.
2.8. Нервные пути и связи в зрительной системе
Из сетчатки зрительная информация по волокнам зрительного нерва устремляется в мозг. Нервы от двух глаз встречаются у основания мозга, где часть волокон переходит на противоположную сторону (зрительный перекрёст, или хиазма). Это обеспечивает каждое полушарие мозга информацией от обоих глаз: в затылочную долю правого полушария поступают сигналы от правых половин каждой сетчатки, а в левое полушарие от левой половины каждой сетчатки (рис. 4.3).
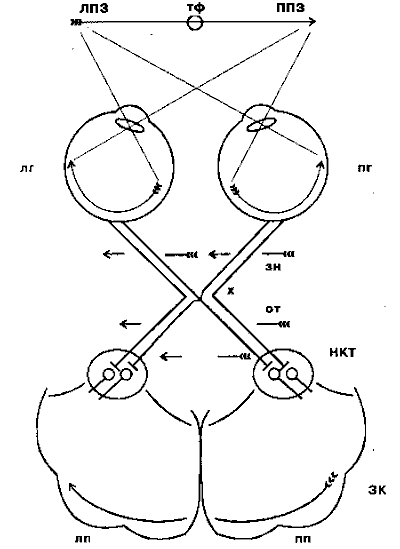
Рис. 4.3. Схема зрительных путей от сетчатки глаза до первичной зрительной коры:
ЛПЗ - левое поле зрения; ППЗ - правое поле зрения; тф - точка фиксации взора; лг - левый глаз; пг - правый глаз; зн - зрительный нерв; х - зрительный перекрёст, или хиазма; от - оптический тракт; НКТ - наружное коленчатое тело; ЗК - зрительная кора; лп - левое полушарие; пп - правое полушарие
После хиазмы зрительные нервы называются оптическими трактами и основное количество их волокон приходит в подкорковый зрительный центр - наружное коленчатoe тело (НКТ). Отсюда зрительные сигналы поступают в первичную проекционную область зрительной коры (стриарная кора, или поле 17 по Бродману). Зрительная кора состоит из ряда полей, каждое из которых обеспечивает свои, специфические функции, получая как прямые, так и опосредованные сигналы от сетчатки и в общем сохраняя её топологию, или ретинотопию (сигналы от соседних участков сетчатки попадают в соседние участки коры).
2.9. Электрическая активность центров зрительной системы
При действии света в рецепторах, а затем и в нейронах сетчатки генерируются электрические потенциалы, отражающие параметры действующего раздражителя (рис. 4.4а, а). Суммарный электрический ответ сетчатки глаза на свет называют электроретинограммой (ЭРГ).

Рис. 4.4. Электроретинограмма (а) и вызванный светом потенциал (ВП) зрительной коры (б):
а,b,с,d на (а) - волны ЭРГ; стрелками указаны моменты включения света. Р 1 - Р 5 - позитивные волны ВП, N 1 - N 5 - негативные волны ВП на (б)
Она может быть зарегистрирована от целого глаза: один электрод помещают на поверхность роговой оболочки, а другой - на кожу лица вблизи глаза (либо на мочку уха). В ЭРГ хорошо отражаются интенсивность, цвет, размер и длительность действия светового раздражителя. Поскольку в ЭРГ отражена активность почти всех клеток сетчатки (кроме ганглиозных клеток), этот показатель широко используется для анализа работы и диагностики заболеваний сетчатки.
Возбуждение ганглиозных клеток сетчатки приводит к тому, что по их аксонам (волокнам зрительного нерва) в мозг устремляются электрические импульсы. Ганглиозная клетка сетчатки это первый в сетчатке нейрон "классического" типа, генерирующий распространяющиеся импульсы. Описано три основных типа ганглиозных клеток: отвечающие на включение света (on - реакция), его выключение (off - реакция) и на то и другое (on-off - реакция). В центре сетчатки рецептивные поля ганглиозных клеток маленькие, а на периферии сетчатки они значительно больше по диаметру. Одновременное возбуждение близко расположенных ганглиозных клеток приводит к их взаимному торможению: ответы каждой клетки становятся меньше, чем при одиночном раздражении. В основе этого эффекта лежит латеральное или боковое торможение (см. гл. 3). Благодаря круглой форме рецептивные поля ганглиозных клеток сетчатки производят так называемое поточечное описание сетчаточного изображения: оно отображается очень тонкой дискретной мозаикой, состоящей из возбужденных нейронов.
Нейроны подкоркового зрительного центра возбуждаются, когда к ним приходят импульсы из сетчатки по волокнам зрительного нерва. Рецептивные поля этих нейронов также круглые, но меньшего размера, чем в сетчатке. Пачки импульсов, генерируемые ими в ответ на вспышку света, короче, чем в сетчатке. На уровне НКТ происходит взаимодействие афферентных сигналов, пришедших из сетчатки, с эфферентными сигналами из зрительной коры, а также из ретикулярной формации от слуховой и других сенсорных систем. Это взаимодействие помогает выделять наиболее существенные компоненты сигнала и, возможно, участвует в организации избирательного зрительного внимания (см. гл. 9).
Импульсные разряды нейронов НКТ по их аксонам поступают в затылочную часть полушарий головного мозга, в которой расположена первичная проекционная область зрительной коры (стриарная кора). Здесь у приматов и человека происходит значительно более специализированная и сложная, чем в сетчатке и в НКТ, переработка информации. Нейроны зрительной коры имеют не круглые, а вытянутые (по горизонтали, вертикали или по диагонали) рецептивные поля (рис. 4.5) небольшого размера [Хьюбел, 1990].

Рис. 4.5 . Рецептивное поле нейрона зрительной коры мозга кошки (А) и ответы этого нейрона на вспыхивающие в рецептивном поле световые полоски разной ориентации (Б). А - плюсами отмечена возбудительная зона рецептивного поля, а минусами - две боковые тормозные зоны. Б - видно, что этот нейрон наиболее сильно реагирует на вертикальную и близкую к ней ориентацию
Благодаря этому они способны выделять из изображения отдельные фрагменты линий с той или иной ориентацией и расположением и избирательно на них реагировать (детекторы ориентаций). В каждом небольшом участке зрительной коры по её глубине сконцентрированы нейроны с одинаковой ориентацией и локализацией рецептивных полей в поле зрения. Они образуют ориентационную колонку нейронов, проходящую вертикально через все слои коры. Колонка - пример функционального объединения корковых нейронов, осуществляющих сходную функцию. Группа соседних ориентационных колонок, нейроны которых имеют перекрывающиеся рецептивные поля, но разные предпочитаемые ориентации, образует так называемую сверхколонку. Как показывают исследования последних лет, функциональное объединение отдалённых друг от друга нейронов зрительной коры может происходить также за счет синхронности их разрядов. Недавно в зрительной коре найдены нейроны с избирательной чувствительностью к крестообразным и угловым фигурам, относящиеся к детекторам 2-гo порядка. Таким образом, начала заполняться "ниша" между описывающими пространственные признаки изображения простыми ориентационными детекторами и детекторами высшего порядка (лица), найденными в височной коре.
В последние годы хорошо исследована так называемая "пространственно-частотная" настройка нейронов зрительной коры [Глезер, 1985; Физиол. зрения, 1992]. Она заключается в том, что многие нейроны избирательно реагируют на появившуюся в их рецептивном поле решётку из светлых и тёмных полос определённой ширины. Так, имеются клетки, чувствительные к решётке из мелких полосок, т.е. к высокой пространственной частоте. Найдены клетки с чувствительностью к разным пространственным частотам. Считается, что это свойство обеспечивает зрительной системе способность выделять из изображения участки с разной текстурой [Глезер, 1985].
Многие нейроны зрительной коры избирательно реагируют на определённые направления движения (дирекциональные детекторы) либо на какой-то цвет (цветооппонентные нейроны), а часть нейронов лучше всего отвечает на относительную удалённость объекта от глаз. Информация о разных признаках зрительных объектов (форма, цвет, движение) обрабатывается параллельно в разных частях зрительной коры.
Для оценки передачи сигналов на разных уровнях зрительной системы часто используют регистрацию суммарных вызванных потенциалов (ВП), которые у человека можно одновременно отводить от сетчатки и от зрительной коры (см. рис. 4.4 б). Сравнение вызванного световой вспышкой ответа сетчатки (ЭРГ) и ВП коры позволяет оценить работу проекционного зрительного пути и установить локализацию патологического процесса в зрительной системе.
2.10. Световая чувствительность
Абсолютная чувствительность зрения . Чтобы возникло зрительное ощущение, свет должен обладать некоторой минимальной (пороговой) энергией. Минимальное количество квантов света, необходимое для возникновения ощущения света в темноте , колеблется от 8 до 47. Одна палочка может быть возбуждена всего 1 квантом света. Таким образом, чувствительность рецепторов сетчатки в наиболее благоприятных условиях световосприятия предельна. Одиночные палочки и колбочки сетчатки различаются по световой чувствительности незначительно. Однако количество фоторецепторов, посылающих сигналы на одну ганглиозную клетку, в центре и на периферии сетчатки различно. Количество колбочек в рецептивном поле в центре сетчатки примерно в 100 раз меньше количества палочек в рецептивном поле на периферии сетчатки. Соответственно и чувствительность палочковой системы в 100 раз выше, чем у колбочковой.
2.11. Зрительная адаптация
При переходе от темноты к свету наступает временное ослепление, а затем чувствительность глаза постепенно снижается. Это приспособление зрительной системы к условиям яркой освещённости называется световой адаптацией. Обратное явление (темновая адаптация) наблюдается, когда из светлого помещения человек переходит в почти не освещённое помещение. В первое время он почти ничего не видит из-за пониженной возбудимости фоторецепторов и зрительных нейронов. Постепенно начинают выявляться контуры предметов, а затем различаются и их детали, так как чувствительность фоторецепторов и зрительных нейронов в темноте постепенно повышается.
Повышение световой чувствительности во время пребывания в темноте происходит неравномерно: в первые 10 мин она увеличивается в десятки раз, а затем, в течение часа - в десятки тысяч раз. Важную роль в этом процессе играет восстановление зрительных пигментов. Так как в темноте чувствительны только палочки, слабо освещённый предмет виден лишь периферическим зрением. Существенную роль в адаптации, помимо зрительных пигментов, играет переключение связей между элементами сетчатки. В темноте площадь возбудительного центра рецептивного поля ганглиозной клетки увеличивается из-за ослабления кольцевого торможения, что приводит к увеличению световой чувствительности. Световая чувствительность глаза зависит и от влияний, идущих со стороны мозга. Освещение одного глаза понижает световую чувствительность неосвещённого глаза. Кроме того, на чувствительность к свету оказывают влияние также звуковые, обонятельные и вкусовые сигналы.
2.12. Дифференциальная чувствительность зрения
Если на освещённую поверхность с яркостью I падает добавочное освещение dI, то, согласно закону Вебера, человек заметит разницу в освещённости только если dI/I = K, где K константа, равная 0,01-0,015. Величину dI/I называют дифференциальным порогом световой чувствительности. Отношение dI/I при разных освещённостях постоянно и означает, что для восприятия разницы в освещённости двух поверхностей одна из них должна быть ярче другой на 1 - 1,5 %.
2.13. Яркостной контраст
Взаимное латеральное торможение зрительных нейронов (см. гл. 3) лежит в основе общего, или глобального яркостного контраста. Так, серая полоска бумаги, лежащая на светлом фоне, кажется темнее такой же полоски, лежащей на тёмном фоне. Это объясняется тем, что светлый фон возбуждает множество нейронов сетчатки, а их возбуждение притормаживает клетки, активированные полоской. Наиболее сильно латеральное торможение действует между близко расположенными нейронами, создавая эффект локального контраста. Происходит кажущееся усиление перепада яркости на границе поверхностей разной освещённости. Этот эффект называют также подчёркиванием контуров, или эффектом Маха: на границе яркого светового поля и более тёмной поверхности можно видеть две дополнительные линии (ещё более яркую линию на границе светлого поля и очень тёмную линию на границе тёмной поверхности).
2.14. Слепящая яркость света
Слишком яркий свет вызывает неприятное ощущение ослепления. Верхняя граница слепящей яркости зависит от адаптации глаза: чем дольше была темновая адаптация, тем меньшая яркость света вызывает ослепление. Если в поле зрения попадают очень яркие (слепящие) объекты, то они ухудшают различение сигналов на значительной части сетчатки (так, на ночной дороге водителей ослепляют фары встречных машин). При тонких работах, связанных с напряжением зрения (длительное чтение, работа на компьютере, сборка мелких деталей), следует пользоваться только рассеянным светом, не ослепляющим глаз.
2.15. Инерция зрения, слитие мельканий, последовательные образы
Зрительное ощущение появляется не мгновенно. Прежде чем возникнет ощущение, в зрительной системе должны произойти многократные преобразования и передача сигналов. Время "инерции зрения", необходимое для возникновения зрительного ощущения, в среднем равно 0,03 - 0,1 с. Следует отметить, что это ощущение также исчезает не сразу после того, как прекратилось раздражение - оно держится ещё некоторое время. Если в темноте водить по воздуху горящей спичкой, то мы увидим светящуюся линию, так как быстро следующие одно за другим световые раздражения сливаются в непрерывное ощущение. Минимальная частота следования световых стимулов (например, вспышек света), при которой происходит объединение отдельных ощущений, называется критической частотой слития мельканий. При средних освещённостях эта частота равна 10-15 вспышкам в 1 с. На этом свойстве зрения основаны кино и телевидение: мы не видим промежутков между отдельными кадрами (24 кадра в 1 с в кино), так как зрительное ощущение от одного кадра ещё длится до появления следующего. Это и обеспечивает иллюзию непрерывности изображения и его движения.
Ощущения, продолжающиеся после прекращения раздражения, называются последовательными образами. Если посмотреть на включённую лампу и закрыть глаза, то она видна ещё в течение некоторого времени. Если же после фиксации взгляда на освещённом предмете перевести взгляд на светлый фон, то некоторое время можно видеть негативное изображение этого предмета, т.е. светлые его части - тёмными, а тёмные - светлыми (отрицательный последовательный образ). Это объясняется тем, что возбуждение от освещённого объекта локально тормозит (адаптирует) определённые участки сетчатки; если после этого перевести взор на равномерно освещённый экран, то его свет сильнее возбудит те участки, которые не были возбуждены ранее.
2.16. Цветовое зрение
Весь видимый нами спектр электромагнитных излучений заключен между коротковолновым (длина волны 400 нм) излучением, которое мы называем фиолетовым цветом, и длинноволновым излучением (длина волны 700 нм), называемым красным цветом. Остальные цвета видимого спектра (синий, зеленый, жёлтый и оранжевый) имеют промежуточные значения длины волны. Смешение лучей всех цветов даёт белый цвет. Он может быть получен и при смешении двух так называемых парных дополнительных цветов: красного и синего, жёлтого и синего. Если произвести смешение трёх основных цветов (красного, зеленого и синего), то могут быть получены любые цвета.
Максимальным признанием пользуется трёхкомпонентная теория Г. Гельмгольца, согласно которой цветовое восприятие обеспечивается тремя типами колбочек с различной цветовой чувствительностью. Одни из них чувствительны к красному цвету, другие - к зеленому, а третьи - к синему. Всякий цвет оказывает воздействие на все три цветоощущающих элемента, но в разной степени. Эта теория прямо подтверждена в опытах, в которых измеряли поглощение излучений с разной длиной волны в одиночных колбочках сетчатки человека.
Частичная цветовая слепота была описана в конце XVIII в. Д. Дальтоном, который сам страдал ею. Поэтому аномалию цветовосприятия обозначили термином "дальтонизм". Дальтонизм встречается у 8% мужчин; его связывают с отсутствием определённых генов в определяющей пол непарной у мужчин X-хромосоме. Для диагностики дальтонизма, важной при профессиональном отборе, используют полихроматические таблицы. Люди, страдающие им, не могут быть полноценными водителями транспорта, так как они могут не различать цвет огней светофоров и дорожных знаков. Существуют три разновидности частичной цветовой слепоты: протанопия, дейтеранопия и тританопия. Каждая из них характеризуется отсутствием восприятия одного из трёх основных цветов. Люди, страдающие протанопией ("краснослепые"), не воспринимают красного цвета, сине-голубые лучи кажутся им бесцветными. Лица, страдающие дейтеранопией ("зеленослепые"), не отличают зеленые цвета от тёмно-красных и голубых. При тританопии (редко встречающейся аномалии цветового зрения) не воспринимаются лучи синего и фиолетового цвета. Все перечисленные виды частичной цветовой слепоты хорошо объясняются трёхкомпонентной теорией. Каждый из них является результатом отсутствия одного из трёх колбочковых цветовоспринимающих веществ.
2.17. Восприятие пространства
Остротой зрения называется максимальная способность различать отдельные детали объектов. Её определяют по наименьшему расстоянию между двумя точками, которые различает глаз, т.е. видит отдельно, а не слитно. Нормальный глаз различает две точки, расстояние между которыми составляет 1 угловую минуту. Максимальную остроту зрения имеет центр сетчатки - жёлтое пятно. К периферии от него острота зрения намного меньше. Острота зрения измеряется при помощи специальных таблиц, которые состоят из нескольких рядов букв или незамкнутых окружностей различной величины. Острота зрения, определённая по таблице, выражается в относительных величинах, причём нормальная острота принимается за единицу. Встречаются люди, обладающие сверхостротой зрения (visus больше 2).
Поле зрения. Если фиксировать взглядом небольшой предмет, то его изображение проецируется на жёлтое пятно сетчатки. В этом случае мы видим предмет центральным зрением. Его угловой размер у человека составляет всего 1,5-2 угловых градуса. Предметы, изображения которых падают на остальные участки сетчатки, воспринимаются периферическим зрением. Пространство, видимое глазом при фиксации взгляда в одной точке, называется полем зрения. Измерение границы поля зрения производят по периметру. Границы поля зрения для бесцветных предметов составляют книзу 70, кверху - 60, внутрь - 60 и кнаружи - 90 градусов. Поля зрения обоих глаз у человека частично совпадают, что имеет большое значение для восприятия глубины пространства. Поля зрения для различных цветов неодинаковы и меньше, чем для чёрно-белых объектов.
Бинокулярное зрение - это зрение двумя глазами. При взгляде на какой-либо предмет у человека с нормальным зрением не возникает ощущения двух предметов, хотя и имеется два изображения на двух сетчатках. Изображение каждой точки этого предмета попадает на так называемые корреспондирующие, или соответственные участки двух сетчаток, и в восприятии человека два изображения сливаются в одно. Если надавить слегка на один глаз сбоку, то начнёт двоиться в глазах, потому что нарушилось соответствие сетчаток. Если же смотреть на близкий предмет, то изображение какой-либо более отдалённой точки попадает на неидентичные (диспаратные) точки двух сетчаток. Диспарация играет большую роль в оценке расстояния и, следовательно, в видении глубины пространства. Человек способен заметить изменение глубины, создающее сдвиг изображения на сетчатках на несколько угловых секунд. Бинокулярное слитие или объединение сигналов от двух сетчаток в единый нервный образ происходит в первичной зрительной коре мозга.
Оценка величины объекта. Величина знакомого предмета оценивается как функция величины его изображения на сетчатке и расстояния предмета от глаз. В случае, когда расстояние до незнакомого предмета оценить трудно, возможны грубые ошибки в определении его величины.
Оценка расстояния. Восприятие глубины пространства и оценка расстояния до объекта возможны как при зрении одним глазом (монокулярное зрение), так и двумя глазами (бинокулярное зрение). Во втором случае оценка расстояния гораздо точнее. Некоторое значение в оценке близких расстояний при монокулярном зрении имеет явление аккомодации. Для оценки расстояния имеет значение также то, что образ знакомого предмета на сетчатке тем больше, чем он ближе.
Роль движения глаз для зрения. При рассматривании любых предметов глаза двигаются. Глазные движения осуществляют 6 мышц, прикреплённых к глазному яблоку. Движение двух глаз совершается одновременно и содружественно. Рассматривая близкие предметы, необходимо сводить (конвергенция), а рассматривая далекие предметы - разводить зрительные оси двух глаз (дивергенция). Важная роль движений глаз для зрения определяется также тем, что для непрерывного получения мозгом зрительной информации необходимо движение изображения на сетчатке. Импульсы в зрительном нерве возникают в момент включения и выключения светового изображения. При длящемся действии света на одни и те же фоторецепторы импульсация в волокнах зрительного нерва быстро прекращается и зрительное ощущение при неподвижных глазах и объектах исчезает через 1-2 с. Если на глаз поставить присоску с крохотным источником света, то человек видит его только в момент включения или выключения, так как этот раздражитель движется вместе с глазом и, следовательно, неподвижен по отношению к сетчатке. Чтобы преодолеть такое приспособление (адаптацию) к неподвижному изображению, глаз при рассматривании любого предмета производит неощущаемые человеком непрерывные скачки (саккады). Вследствие каждого скачка изображение на сетчатке смещается с одних фоторецепторов на другие, вновь вызывая импульсацию ганглиозных клеток. Продолжительность каждого скачка равна сотым долям секунды, а амплитуда его не превышает 20 угловых градусов. Чем сложнее рассматриваемый объект, тем сложнее траектория движения глаз. Они как бы "прослеживают" контуры изображения (рис. 4.6), задерживаясь на наиболее информативных его участках (например, в лице это глаза). Кроме скачков, глаза непрерывно мелко дрожат и дрейфуют (медленно смещаются с точки фиксации взора). Эти движения также очень важны для зрительного восприятия.

Рис. 4.6. Траектория движения глаз (Б) при осматривании изображения Нефертити (А)


