Kahepaiksete aju areng. Kahepaiksed ehk kahepaiksed
Kahepaiksed ehk kahepaiksed täiskasvanuna on tavaliselt maismaaloomad, kuid siiski on nad tihedalt seotud veekeskkonnaga ning nende vastsed elavad pidevalt vees. Järelikult vene ja kreeka keel (amfiibid - juhtiv topeltelu) nimed peegeldavad nende selgroogsete peamist tunnust. Kahepaiksed tekkisid, nagu eespool mainitud, väikestes mageveekogudes elanud devoni sagaruimelistest kaladest, kes roomasid oma lihakate paariuimede abil kaldale.
Väline hoone. Keha (joonis 147) koosneb peast, torsost, esi- ja tagaosast, mis on paaris tükeldatud. Jäsemed koosnevad kolmest osast: eesmised - õlast, küünarvarrest ja käest, tagumised - reiest, säärest ja labajalast. Vaid vähesel osal tänapäeva kahepaiksetest on saba (sabade järjekord – vesilikud, salamandrid jne). Kahepaiksete suurima rühma - anuraanide (konnad, kärnkonnad jt) täiskasvanud vormidel on see vähenenud viimaste kohanemise tõttu maismaal hüpates liikumisega, kuid säilib nende vastsetes - vees elavates kullestes. Mõnel poolmaa-aluse eluviisiga liigil (jaladeta järjekord ehk caecilians) olid jäsemed ja saba vähenenud.
Pea on kehaga liikuvalt liigendatud, kuigi selle liikumine on väga piiratud ja kael puudub. Tükeldatud jäsemed ning liigutatav ühendus pea ja keha vahel on iseloomulikud tunnused maismaa selgroogsed, kalades need puuduvad. Maapealsete vormide keha on dorso-ventraalses suunas lapik, kaladel (ujumisega kohanemise tõttu) aga reeglina külgsuunas kokkusurutud. Vees elavatel kahepaiksetel läheneb kehakuju kala omale. Keha suurus on vahemikus 2–160 cm (Jaapani salamander); keskmiselt on kahepaiksed väiksemad kui teised maismaaloomad. Nahk paljas näärmete rikas, eraldunud paljudes kohtades lihastest nahaaluste lümfiõõnsuste olemasolu tõttu. See on varustatud suure hulgaga veresooned ja täidab ka hingamisfunktsiooni (vt allpool). Teatud tüüpi tühjenemise korral naha näärmed mürgine. Nahavärv on väga mitmekesine.
Närvisüsteem. Seoses kahepaiksete kohanemisega maismaaeluga ja eriti seoses liikumisloomuse radikaalse muutumisega on närvisüsteem päris palju muutunud. Kahepaiksete eesaju (vt joon. 133, B) on keskmisest suurem; kalades täheldatakse reeglina vastupidist suhet. Seda seletatakse asjaoluga, et kaladel on funktsioonid eesaju on seotud ainult haistmisstiimulite tajumisega kahepaiksetel, hakkab ta osalema erinevate kehafunktsioonide koordineerimisel ja selle pinnakihis tekivad ajukoore alged (ikka väga nõrgad), millesse on koondunud närvirakud. Samas tuleb märkida, et eesajus on hästi arenenud haistmissagarad. Kahepaiksete väikeaju on erinevalt kaladest väga halvasti arenenud. Kalad liiguvad pidevalt ja nende kehaasend on ebastabiilne, samas kui kahepaiksed, kes toetuvad jalgadele, on üsna stabiilses asendis. saidid selgroog, nendes kohtades, kus närvid sellest lahkuvad, lähevad jalalihastesse, mis teevad palju rohkem tööd kui kalade paariuimede lihased, paksenevad ning nendega on ühendatud õlavarre- ja nimmepõimikud. Perifeerne närvisüsteem on lihaste diferentseerumise (vt allpool) ja pikkade liigeste jäsemete ilmnemise tõttu suuresti muutunud.

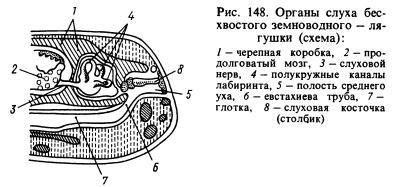
Meeleelunditest on kõige olulisemad muutused läbi teinud kuulmisorgan. Helilainete ülekanne veekeskkonnast loomade kudedesse, mis on samuti veega küllastunud ja millel on ligikaudu samad akustilised omadused kui vees, toimub palju paremini kui õhust. Sisse rändavad helilained õhukeskkond, peegelduvad peaaegu täielikult looma pinnalt ja ainult umbes 1% nende lainete energiast tungib tema kehasse. Sellega seoses arenesid lisaks labürindile välja kahepaiksed või sisekõrv, kuulmisorgani uus osa on keskkõrv. See on (joonis 148) väike õhuga täidetud õõnsus, mis suhtleb suuõõnega läbi Eustachia toru ja on väljastpoolt suletud õhukese elastse kuulmekilega. Keskkõrvas on kuulmisplaat (või sammas), mis ühest otsast toetub vastu kuulmekile, teisest otsast vastu kilega kaetud akent, mis viib koljuõõnde, kus on perilümfiga ümbritsetud labürint. Rõhk keskkõrva sees on võrdne atmosfäärirõhuga ja kuulmekile võib vibreerida heli õhulainete mõjul, mille mõju kandub edasi läbi kuulmisluu ja perilümfi labürindi seintele ning on tajutav kuulmisnärvi otste kaudu. Keskkõrva õõnsus moodustati esimesest lõpusepilust ja sammas moodustati pilu lähedal paiknevast hüomandibulaarsest luust (hüomandibulaarluust), mis riputas kolju vistseraalse osa ajju, kus labürint paiknes lõhe taga. kõrva luud.
Silmad on kaetud liikuvate silmalaugudega, mis kaitsevad nägemisorganeid kuivamise ja ummistumise eest. Tänu sarvkesta ja läätse kuju muutustele näevad kahepaiksed kaladest kaugemale. Kahepaiksed tajuvad väikseid temperatuurimuutusi hästi. Nad on mõjude suhtes tundlikud erinevaid aineid, lahustatud vees. Nende haistmisorgan reageerib gaasiliste ainete põhjustatud ärritustele. Seega on kahepaiksete meeleelundid seoses maismaal elamisele üleminekuga läbi teinud mitmeid muutusi. Pidevalt vees elavatel vastsetel ja täiskasvanud loomadel on nagu kaladel ka külgjoonorganid.
Kahepaikseid iseloomustavad üsna keerulised instinktiivsed tegevused, eriti pesitsusperioodil. Näiteks Venemaal Lääne-Ukrainas elav isane ämmaemand kärnkonn mähib oma tagajäsemete ümber munade “nöörid” ja peidab end kaldale eraldatud kohtadesse, kuni kullesed arenevad. 17-18 päeva pärast naaseb isane vette, kus kullesed kooruvad. See on omamoodi instinkt järglaste kaitsmiseks. Paljudel troopilistel sabata kahepaiksetel on teada veelgi keerulisemad instinktid. Kahepaiksetel on ka konditsioneeritud refleksid, kuid need arenevad suurte raskustega.
Mootorsüsteem ja luustik. Lihassüsteem seoses erinevaid seadmeid elule maismaal (maa-tüüpi jäsemete areng, liikuva liigese tekkimine pea ja keha vahel jne) läbis radikaalseid muutusi, kuigi see säilitas palju kaladele omaseid jooni. Kalade lihassüsteem on väga ühtlane ja koosneb peamiselt sarnastest külgmistest lihassegmentidest. Kahepaiksete lihassüsteem on muutunud diferentseeritumaks, koosnedes mitmesugustest lihastest (joonis 149). Kahepaiksed panid aluse lihassüsteemile, mis hiljem arenes ja muutus keerukamaks tõeliste maismaaselgroogsete – roomajate, lindude ja imetajate puhul. See kehtib ka luustiku kohta.
Kahepaiksete koljus on palju kõhrelisi elemente, mis on ilmselt seletatav poolmaisest eluviisist tuleneva vajadusega kergendada kehakaalu. Kolju sisaldab palju kõrgemate kalade kolju kirjelduses loetletud luid, sealhulgas ainult kaladele ja kahepaiksetele iseloomulikku parasfenoidi (joon. 150). Kuna hüomandibulaarsest luust on saanud kuulmisluu, siis mängib ripatsi rolli nelinurkne luu. Lõpuseaparaadi kadumise tõttu täiskasvanueas vähenevad lõpusekaared ja säilivad vaid nende muudetud jäänused. Hüoidkaar muutub suuresti ja on osaliselt vähenenud. Kahepaiksete kolju on väga lai, mis on osaliselt tingitud nende hingamise iseärasustest. Alumine lõualuu, nagu ka luukaladel, koosneb mitmest luust.
Lülisammas (joonis 150) on sabata loomadel väga lühike ja lõpeb pika luuga - urostiiliga, mis on moodustatud sabalülide algetest. Sabaga kahepaiksetel koosneb lülisamba sabaosa paljudest selgroolülidest. Nendel kahepaiksetel on sabal liikumisel oluline roll: vees kasutatakse seda ujumiseks, maismaal tasakaalu säilitamiseks. Roided on halvasti arenenud (sabakujulistel kahepaiksetel) või vähenenud ning nende jäänused on sulanud selgroolülide põikprotsessidega (teistel kahepaiksetel olid ribid). Nende vähenemist tänapäevastes vormides seletatakse vajadusega kergendada nende selgroogsete kehamassi (mis kasvas veekeskkonnast õhku üleminekul oluliselt), kes pole veel maismaal liikumiseks piisavalt kohanenud. Roiete vähenemise tõttu kahepaiksetel puuduvad rind. Esimene selgroolüli on üles ehitatud teisiti kui kaladel: tal on kaks glenoidne lohk liigendamiseks kolju kahe kuklaluu kondüüliga, mille tõttu kahepaiksete pea liikus.
Esijäseme skelett (joon. 150) koosneb õlavarreluu, kaks küünarvarre luud - raadius ja küünarluu, randme luud, kämblaluud ja sõrmede falangid. Tagajäseme luustik (joonis 150) koosneb reiest, sääre kahest luust - sääreluust ja pindluust, tarsaalluust, pöialuudest ja sõrmede falangetest. Järelikult on mõlema jäsemepaari struktuuri sarnasus, hoolimata nende funktsioonide mõningatest erinevustest, väga suur. Esialgu olid esi- ja tagajalad viievarbalised, tänapäeva kahepaiksetel võib olla vähem varbaid. Ujumiseks kasutatakse ka paljude sabata kahepaiksete tagajäsemeid ja seetõttu on need piklikud ning sõrmi ühendavad ujumismembraanid.
Jäsemete vööd on palju paremini arenenud kui kaladel. Õlavööde koosneb luu- ja kõhreelementidest: abaluu, rangluu, vareseluu (korakoid) jne (joon. 150). Randluud ja korakoid on ühendatud rinnakuga, mis sisaldab ka luu- ja kõhreelemente. Õlavöötmega liigendub õlavarreluu pea. Jäsemete tagumine vöö ehk vaagnaluu koosneb kolmest luust: ilium, häbemeluu ja ischium (joon. 150). Nendest luudest moodustunud suur astabulum on mõeldud reieluu peaga liigendamiseks. Vaagen on ühendatud ühe selgroolüliga - ristluuga, tänu millele said tagajalad erinevalt kalade kõhuuimedest üsna tugeva toe.
Vereringe. Vees elavate ja lõpustega hingavate kahepaiksete vastsetel on vereringesüsteem põhimõtteliselt sarnane kalade vereringesüsteemiga, kuid maismaa eluviisi juhtivatel täiskasvanud loomadel muutub see oluliselt, kuna lõpusehingamine asendub kopsuhingamisega. suurenenud nahahingamine ning maismaaloomade jäsemete areng ja muud keha muutused. Süda (vt. Joon. 134, B, 151) koosneb kolmest kambrist: parem- ja vasakpoolne koda ning üks vatsake. Väljub viimase paremalt küljelt arterioosne koonus(see esines ka kaladel, kahepaiksete esivanematel), millest pärinevad neli paari artereid: esimene paar - unearterid , verekandjad pähe, teine ja kolmas paar on veresooned, mis ühenduvad, moodustades keha suurima anuma - aordi, mille oksad on suunatud erinevatesse kehaosadesse, neljas paar - kopsuarterid, mis jagunevad seejärel iseseisvateks naha- ja kopsuarteriteks.
Kopsudest voolab hapnikurikas veri kopsuveenide kaudu vasakusse aatriumisse ja küllastunud veri kõikidesse kehaosadesse süsinikdioksiid, keha eesmises osas voolab see eesmisse õõnesveeni ja keha tagumises osas tagumisse õõnesveeni (joon. 152). Mõlemad õõnesveenid tühjenevad sisse venoosne siinus, kust suubub veri (süsihappegaasiga küllastunud). parem aatrium. Mõlemast kodadest siseneb veri südame üksikusse vatsakesse. Vatsakese sisepinnal on süvendid ja seetõttu ei ole selles verel aega täielikult seguneda: vasakus osas on hapnikuga küllastunud veri, paremas osas süsihappegaasiga küllastunud veri ja keskmises osas. see on segatud. Kuna arteriooskoonus algab parem pool vatsakesesse, siis esimene osa verest, mis sinna (st arterikoonusse) siseneb, on venoosne, suunatakse see kõige tagumatesse arteritesse - kopsuarteritesse.
Seguveri, mis seejärel voolab aordi moodustavatesse arteritesse ja viimase harude kaudu kõikidesse kehaosadesse. Hapnikuga varustatud veri vatsakese vasakust küljest saadetakse unearteritesse. Sellele peame lisama, et naha hapnikuga küllastunud veri siseneb, nagu eespool märgitud, eesmise õõnesveeni ja venoosse siinuse kaudu paremasse aatriumisse ja seega lahjendab seal olevat verd. venoosne veri, seejärel surutakse aordi moodustavatesse anumatesse. Järelikult saavad erinevad kehaosad tänu ülalkirjeldatud seadmetele ja ka teistele siin kirjeldamata seadmetele ebavõrdselt hapnikuga küllastunud verd. Joonisel fig. 152 näitab peamist arteriaalset ja venoossed veresooned kahepaiksed.
Kahepaiksetel on jäsemete tugeva arengu ja suurema keha dissektsiooni tõttu kui kaladel veresoonte võrgustik oluliselt muutunud. Ilmus palju uusi laevu, mis kaladel puudusid, ja tekkis maismaaselgroogsetele iseloomulik laevade süsteem. Samas tuleb meeles pidada, et kahepaiksete vereringesüsteem on palju lihtsam kui kõrgematel selgroogsetel.
Hingamissüsteem. Peaaegu kõigil kahepaiksetel on kopsud (vt joon. 151; 153). Need elundid on endiselt väga lihtsa ehitusega ja õhukese seinaga kotikesed, mille seintes hargneb üsna tihe veresoonte võrgustik. Sest sisemine sein Kopsud on peaaegu siledad, siis on nende pind suhteliselt väike. Hingetoru on peaaegu välja arenemata ja kopsud on ühendatud otse kõriga. Kuna kahepaiksetel ei ole rindkere (vt eespool), tagab hingamistegevuse suuõõne lihaste töö. Sissehingamine toimub järgmiselt. Avatud ninasõõrmetega (mis erinevalt kalade ninasõõrmetest on läbivad, st lisaks välistele ninasõõrmetele on ka sisemised ninasõõrmed - choanae) Ja suletud suu Suure suuõõne põhi tõmmatakse tagasi ja õhk siseneb sinna. Seejärel suletakse ninasõõrmed spetsiaalsete klappidega, tõstetakse suupõhja ja surutakse õhk kopsudesse. Väljahingamine toimub kõhulihaste kokkutõmbumise tagajärjel.
Kahepaiksed saavad märkimisväärses koguses hapnikku läbi naha ja suuõõne limaskestade. Mõnel salamandriliigil pole üldse kopse ja kogu gaasivahetus toimub läbi naha. Hingamisfunktsioone saab nahk aga täita vaid siis, kui see on niiske. Seetõttu on kahepaiksetel võimatu elada tingimustes, kus õhuniiskus on nende jaoks ebapiisav. Vees elavad vastsed hingavad läbi lõpuste (kõigepealt välised, siis sisemised) ja naha. Mõnel pidevalt vees elaval sabaga kahepaiksel on lõpused, mis püsivad kogu elu. Seega on kahepaiksed hingamisviiside poolest endiselt kaladele lähedased.
Eritussüsteem. Neerud (vt. joon. 136, A, B; joon. 151), nagu kaladel, on tüve. Wolffi kanalid tühjenevad kloaaki. Seal avaneb põis, kuhu koguneb uriin. Dissimilatsioonitoodete eemaldamine toimub ka naha ja kopsude kaudu.
Seedeelundkond. Suuõõs on väga lai. Paljudel liikidel (peamiselt sabaga kahepaiksetel) on palju väikeseid, ühtlaseid, ürgselt paigutatud hambaid, mis istuvad lõualuudel, vomeril, palatinal ja muudel luudel ning on mõeldud ainult saagi hoidmiseks. Enamikul liikidel (peamiselt sabata kahepaiksetel) on hambad osaliselt või täielikult vähenenud, kuid nende keel areneb tugevasti. Viimane konnadel on kinnitatud esiotsa ja selle saab saagi püüdmiseks tagumise otsaga kaugele ette visata. See on väga kleepuv ja selle funktsiooni täitmiseks hästi kohandatud. Pidevalt vees elavatel liikidel on keel tavaliselt vähenenud. Selliste kahepaiksete saakloomade püüdmine toimub lõualuude abil.
Seedetoru (vt joonis 151) on suhteliselt lühike ja koosneb neelust, söögitorust, maost, peensoolest ja väga väikesest pärasoolest (käärsoolest). Pärasoole tagumine osa on kloaak; selle kaudu eritub lisaks väljaheitele ka uriin ja seksuaalproduktid. Süljenäärmed, mis kaladel puuduvad, voolavad suuõõnde. Nende näärmete sekretsioon on mõeldud peamiselt toidu niisutamiseks. Süljenäärmed on vees elavatel liikidel väga halvasti arenenud, maapealsetel aga palju paremini. Maks on suur; kõhunääre on hästi määratletud. Täiskasvanud kahepaiksete toit on peamiselt loomne (putukad, väikesed selgroogsed jne). Sabata kahepaiksete kullesed on enamasti taimtoidulised.

Paljundamine. Meeste sugunäärmed (munandid) asuvad neerude lähedal (vt joon. 151, B). Nende kanalid avanevad neerude eesmise osa tuubulitesse (vt. joon. 136, A) ja seeme eritub sarnaselt uriiniga Wolffi kanalite kaudu. Naiste sugunäärmed (munasarjad) kasvavad kudemisperioodil tugevasti. Munad väljuvad väga pikkade Mülleri kanalite kaudu (vt joon. 136, B). Viimastel puudub otsene seos munasarjadega ning valmivad munad sisenevad kehaõõne kaudu Mülleri kanalite lehtritesse.
Enamasti toimub väetamine vees. Paljudel kahepaiksetel eelneb sellele isase ja emase lähenemine: isane haarab emase selja tagant kinni, surub oma esijäsemed talle peale. kõhu seina ja see hõlbustab munade vette laskmist, mille ta kohe viljastab. Seega toimub seksuaalvahekorras viljastumine väljaspool naise keha. Vähemustel liikidel (näiteks vesilased) eraldab isaslind seemne spetsiaalses kotis (spermatofoor), mille emane kloaagi servadega kohe kinni püüab. Sel juhul seksuaalvahekorda ei toimu, vaid viljastumine on sisemine. Lõpuks sisestab isasloom mõne liigi puhul seemne emase kloaaki, kasutades oma väljaulatuvat kloaaki.
Paljudel liikidel on seksuaalne dimorfism hästi väljendunud (värvus, esijalgade struktuur, millega isased emasloomi hoiavad, ja muud omadused). Paljude liikide isased võivad teha väga valjud helid tänu nende helide võimendamisele häälekottide – resonaatorite poolt.
Areng. Kahepaiksete areng toimub tavaliselt vees. Viljastatud munadest arenevad vastsed (kullesed), millel on kalataoline kuju. Nad hingavad läbi lõpuste ja sisemine struktuur meenutab kala. Kasvuperioodil toimub kulleste transformatsioon (metamorfoos): esmalt kasvavad tagajalad, seejärel esijalad, lõpused ja saba atroofia (sabata loomadel), kopsud, Suured muutused esinevad vereringesüsteemis jne.
Päritolu. Kahepaiksed, nagu eespool selgitatud (lk 296), põlvnesid laba-uimkaladest. Iidsete laba-uimeliste kalade paarisuimed, millest arenesid maismaaselgroogsete liigendjäsemed, olid lühikesed ja laiad, sisaldasid palju väikeseid, liigestega mitteühendatud luuelemente, mis paiknesid mitmes (vähemalt kaheksas) põikreas. Vööd, millele uimed toetusid, olid suhteliselt halvasti arenenud (eriti vaagnavöö). Uimede muutumise tõttu maapealseteks jäsemeteks toimusid skeletis olulised muutused.
Esiteks vähendati paljusid luuelemente: esimestes proksimaalsetes ridades oli alles ainult üks luu, esijalas - õlg, taga - reieluu; teistes ridades - kaks luud, esijalas - raadius ja küünarluu, taga - sääreluu ja pindluu; kahes järgmises reas jäid alles viis luud, mis moodustasid esijalas randme ja tagumise jala randme; V järgmine ridaülejäänud viis luud kuulusid esijala kämblasse ja tagumise jala pöialuudesse; ülejäänud kolmest viie luuga reast said sõrmede falangid. Luude arvu vähendamine aitas kaasa jalgade tugevuse suurenemisele.
Teiseks pikendati oluliselt kahe esimese rea luid (st õlg ja käsivars, reie ja sääreosa), mis oli liikumiskiiruse suurendamiseks väga oluline.
Kolmandaks arenesid loetletud luude vahel liigesed ehk jäsemed liigendusid, mis on nende töö kõige olulisem tingimus.
Neljandaks on tugevdatud jäsemete vööd (vt vööde kirjeldust ülalt). Paralleelselt need muudatused on toimunud põhjalikud muutused närvi-, lihas- ja veresoonte süsteemid jalad Muutused teistes elundisüsteemides, mis toimusid uimeliste kalade kahepaikseteks muutumisel, on kirjeldatud aastal. üldised omadused viimane.
Kõige iidsemad kahepaiksed olid stegotsefaalid (joon. 154), keda oli süsinikuperioodil arvukalt ja kes surid lõplikult välja triiase ajal. Nad elasid veehoidlate kallastel ja veetsid palju aega vees. Nende loomade pea oli kaetud naastudega, mis selgitab nende nime (stegocephals - kaetud peaga). Nende luustikul oli palju primitiivseid jooni: jalgade luuelemendid olid väikesed ja veidi erineva suurusega, selgroolülid olid kaksiknõgusad, jäsemete vööd olid halvasti arenenud jne. Nendest tekkisid tänapäevased kahepaiksete rühmad.
Aju konnasid, nagu ka teisi kahepaikseid, iseloomustavad kaladega võrreldes järgmised omadused:
A) progressiivne areng aju, mis väljendub paarispoolkerade eraldumises pikilõhega ja iidse ajukoore (archipalliumi) halli aine arengus aju katuses;
b) väikeaju nõrk areng;
c) aju kõverate nõrk väljendus, mille tõttu vahepealsed ja keskmised osakonnadülalt selgelt nähtav.
Teemant aju(rombencephalon)
Medulla oblongata (müelencephalon, medulla oblongata), millesse seljaaju kraniaalselt läbib, erineb see viimasest suurema laiuse ja tagumiste kraniaalnärvide suurte juurte lahkumise poolest külgpindadest. Medulla oblongata seljapinnal on rombikujuline lohk (fossa rhomboidea), vastutulelik neljas ajuvatsake (ventriculus quartus). Pealt on see kaetud õhukesega veresoonte kork, mis kustutatakse koos ajukelme. Ventraalne lõhe, seljaaju ventraalse lõhe jätk, kulgeb mööda pikliku medulla ventraalset pinda. Medulla oblongata sisaldab kahte paari nööre (kiudude kimpu): alumine paar, mis on eraldatud ventraalse lõhega, on motoorne, ülemine paar on sensoorsed. Medulla oblongata sisaldab lõualuu ja keelealuse aparatuuri keskusi, kuulmisorganit, samuti seede- ja hingamissüsteeme.
Väikeaju asub rombikujulise lohu ees kõrge põikharja kujul selle esiseina väljakasvuna. Väikeaju väiksuse määrab kahepaiksete väike ja ühtlane liikuvus – tegelikult koosneb see kahest väikesest osast, mis on tihedalt seotud medulla oblongata akustiliste keskustega (need osad säilivad imetajatel kui väikeaju fragmendid (flokulid)). Väikeaju keha – koordinatsioonikeskus teiste ajuosadega – on väga halvasti arenenud.
Keskaju (mesencephalon) seljaküljelt vaadatuna esindab seda kaks tüüpilist optilised labad (lobus opticus s. tectum opticus), millel on paaris munakujulised tõusud, mis moodustavad keskaju ülemise ja külgmise osa. Optiliste sagarate katuse moodustab hall aine - mitu kihti närvirakud. Kahepaiksete tektum on aju kõige olulisem osa. Optilised lobud sisaldavad õõnsusi, mis on külgmised oksad aju (Sylvii) akvedukt (aquaeductus cerebri (Sylvii), mis ühendab neljanda ajuvatsakese kolmandaga.
Keskaju põhja moodustavad paksud kimbud närvikiud - jalad suur aju(cruri cerebri),ühendab eesaju pikliku medulla ja seljaajuga.
Eesaju(prosentsefalon) koosneb vahe- ja telentsefalon, lamades seerias.
Diencephalon pealtpoolt nähtav rombina, külgedele suunatud teravate nurkadega.
Vahelihase osad asuvad vertikaalselt paikneva laia lõhe ümber kolmas ajuvatsake (ventriculus tertius). Vatsakese seinte külgmine paksenemine - visuaalsed täpid või talamus. Kaladel ja kahepaiksetel on talamus teisejärgulise tähtsusega (sensoorsete ja motoorsete keskustena). Kolmanda ajuvatsakese – epitalamuse ehk epitalamuse – membraanne katus ei sisalda neuroneid. See sisaldab ülemist medullaarnääre - käbinääre (epifiis). Kahepaiksetel toimib käbinääre juba näärmena, kuid pole veel kaotanud parietaalse nägemisorgani tunnuseid. Epifüüsi ees on vahekeha kaetud membraanse katusega, mis suu kaudu pöördub sissepoole ja läheb eesmisse koroidpõimikusse (kolmanda vatsakese koroidpõimikusse) ja seejärel vahepea otsplaadisse. Altpoolt vatsake kitseneb, moodustub hüpofüüsi lehter (infundibulum), alumine medullaarnääre on selle külge kinnitatud kaudoventraalselt - hüpofüüs (hüpofüüs). Ees, terminali põhja ja aju vahepealsete osade vahelisel piiril, on rist nägemisnärvid(chiasma nervorum opticorum). Kahepaiksetel ei jää enamus nägemisnärvi kiude vaheajusse, vaid lähevad kaugemale – keskaju katusele.
Teleencephalon selle pikkus on peaaegu võrdne kõigi teiste ajuosade pikkusega. See koosneb kahest osast: haistmisajust ja kahest poolkerast, mis on üksteisest eraldatud sagitaalne (noolekujuline) lõhe (fissura sagittalis).
Teletsefaloni poolkerad (haemispherium cerebri) hõivavad kaks kolmandikku telentsefaloni tagumisest osast ja ripuvad vahelihase esiosa kohal, kattes seda osaliselt. Poolkerade sees on õõnsused - külgmised ajuvatsakesed (ventriculi lateralis), kaudaalselt suhelda kolmanda vatsakesega. Kahepaiksete ajupoolkerade hallaines võib eristada kolme piirkonda: dorsomediaalselt on vana ajukoor ehk hipokampus (archipallium, s. hippocampus), külgsuunas - iidne koor(paleopallium) ja ventrolateraalselt - basaalganglionid, vastavad striata (corpora striata) imetajad. Striatum ja vähemal määral hipokampus on korrelatsioonikeskused, viimane on seotud haistmisfunktsioon. Iidne koor on eranditult lõhnaanalüsaator. Poolkerade ventraalsel pinnal on märgatavad sooned, mis eraldavad juttkeha iidsest ajukoorest.
Haistmisaju (rhinencephalon) hõivab telentsefaloni esiosa ja moodustab lõhnasagar (lobus olfactorius), joodetud keskelt omavahel. Need on poolkeradest külgmiselt eraldatud marginaalse lohuga. Haistmissagarad sisaldavad eesmiselt haistmisnärve.
Konna ajust ulatub välja 10 paari kraniaalnärvid. Nende moodustumine, hargnemine ja innervatsioonitsoon ei erine põhimõtteliselt imetajate omast
Lihassüsteem. Ta erineb sellistest kaladest peamiselt jäsemete lihaste suurema arengu ja kehatüve lihaste suurema diferentseerumise poolest, mis koosneb üksikute lihaste keerukast süsteemist. Selle tulemusena on esmane lihase segmentatsioon häiritud, kuigi mõnes kõhu- ja seljalihases jääb see siiski üsna selgeks.
Närvisüsteem. Kahepaiksete aju erineb kalade ajust peamiselt eesaju suurema arengu, selle poolkerade täieliku eraldumise ja vähearenenud väikeaju poolest, mis kujutab endast vaid neljanda vatsakese eesmist osa kattev väikest närviainest ribi. Eesaju areng ei väljendu mitte ainult selle suurenemises ja diferentseerumises, vaid ka selles, et lisaks külgvatsakeste põhjale sisaldavad nende küljed ja katus närviaine, see tähendab, et kahepaiksetel tekib tõeline ajuvõlv - arhipallium (tänapäeva kaladel on kopsukaladel arhipallium). Haistmissagarad on poolkeradest vaid nõrgalt piiritletud. Vahepealset osa katavad ülalt vaid veidi naaberlõigud. Parietaalelund on kinnitatud selle katusele ja põhjast ulatub hästi määratletud lehter, mille külge on kinnitatud hüpofüüs. Kuigi keskaju on märkimisväärne osa, on see suhteliselt väiksem kui kaladel. Väikeaju alaareng, nagu kopsukaladelgi, on seotud kehaliigutuste lihtsusega: kahepaiksed on üldiselt istuvad loomad, kuid neil, kes nagu konnadki suudavad teha kiireid liigutusi, piirduvad nad hüppamisega, s.t väga lihtsate liigutustega. Ajust, nagu kondine kala, tekib ainult 10 paari peanärve; XII paar (hüpoglossaalne närv) ulatub väljapoole kolju ja XI paar (lisanärv) ei ole üldse välja arenenud.

. I - ülemine; II - põhi; III - külg; IV - pikisuunas (vastavalt Parkerile):
1 - eesaju poolkerad, 2 - haistmissagara, 3 - haistmisnärv, 4 - vaheaju poolkerad, 5 - visuaalne kiasm, 6 — lehter, 7 - ajuripats, 8 - keskaju, 9 - väikeaju, 10 - medulla oblongata, 11 - neljas vatsake, 12 - seljaaju, 13 - kolmas vatsake, 14 - Sylviuse akvedukt,
III - X - peanärvid, XII - hüpoglossaalne närv

, skeem (Gregory järgi):
1 - kolju, 2 - medulla oblongata, 3 - kuulmisnärv, 4 - poolringikujulised kanalid, 5 - keskkõrvaõõs, 6 - Eustachia toru, 7 - neelu, 8 - kõrv, 9 - kuulmekile
Konnas on 10 paari tõelisi seljaajunärve. Formeerimisel osalevad kolm esipaari brahiaalne põimik, innerveerivad esijäsemeid ja neli tagumist paari – nimme-ristluupõimiku moodustamisel, innerveerivad tagajäsemeid.
Konna sümpaatiline närvisüsteem, nagu ka kõigil kahepaiksetel, on väga hästi arenenud ja seda esindavad peamiselt kaks närvitüve, mis ulatuvad mõlemal pool selgroogu ja on moodustatud närviganglionide ahelast, mis on omavahel ühendatud nööride ja nööride kaudu. ühendatud seljaaju närvidega.
Veel huvitavaid artikleid
Kalades aju tervikuna on väike. See on halvasti arenenud eesmine osa. Eesaju pole poolkeradeks jagatud. Selle katus on õhuke, koosneb ainult epiteelirakkudest ja ei sisalda närvikudet. Eesaju põhi sisaldab juttkeha ja sellest ulatuvad välja haistmissagarad. Funktsionaalselt on eesaju kõrgeim haistmiskeskus.
Diencephalonis, millega on ühendatud käbinääre ja hüpofüüs, asub hüpotalamus, mis on endokriinsüsteemi keskne organ. Kalade keskaju on kõige arenenum. See koosneb kahest poolkerast ja on kõrgeim visuaalne keskus. Lisaks esindab see aju kõrgeimat integreerivat osa. Tagaajus on väikeaju, mis reguleerib liigutuste koordineerimist. See on väga hästi arenenud seoses kalade liikumisega kolmemõõtmelises ruumis. Medulla oblongata tagab side aju kõrgemate osade ja seljaaju vahel ning sisaldab hingamis- ja vereringekeskusi. Seda tüüpi aju, mille kõrgeimaks funktsioonide integratsioonikeskuseks on keskaju, nimetatakse ihtüopsid.
Kahepaiksetel aju on ka ihtüopsid. Nende eesaju on aga suur ja jagatud poolkeradeks. Selle katus koosneb närvirakkudest, mille protsessid asuvad pinnal. Sarnaselt kaladele ulatub keskaju suur suurus, mis esindab ka kõrgeimat integreerivat keskust ja nägemiskeskust. Väikeaju on liigutuste primitiivse olemuse tõttu mõnevõrra vähenenud. Maapealse eksisteerimise tingimused roomajad nõuavad aju keerukamat morfofunktsionaalset korraldust. Eesaju on teistega võrreldes suurim osa. Selles on eriti arenenud juttkeha. Kõrgeima integreeriva keskuse funktsioonid lähevad neile üle. Esimest korda ilmuvad katuse pinnale väga primitiivse struktuuriga kooresaared iidne - arhikorteks. Keskaju kaotab oma tähtsuse juhtiva sektsioonina ja selle suhteline suurus väheneb. Väikeaju on roomajate liigutuste keerukuse ja mitmekesisuse tõttu kõrgelt arenenud. Seda tüüpi aju, mille juhtivat sektsiooni esindavad eesaju striataalsed kehad, nimetatakse sauropsid.
Imetajatel - imetajad aju tüüp. Seda iseloomustab eesaju tugev areng tänu ajukoorele, mis areneb roomaja ajukoore väikese saare baasil ja muutub aju integreerivaks keskuseks. See sisaldab kõrgeimaid nägemis-, kuulmis-, puutekeskusi, mootori analüsaatorid, samuti kõrgema närvisüsteemi aktiivsuse keskused. Kooril on väga keeruline struktuur ja kutsutakse uus koor - neokorteks. See ei sisalda mitte ainult neuronite kehasid, vaid ka selle erinevaid osi ühendavaid assotsiatiivseid kiude. Iseloomulik on ka mõlema poolkera vahelise komissiooni olemasolu, milles paiknevad neid omavahel ühendavad kiud. Sidefalon, nagu ka teised klassid, hõlmab hüpotalamust, hüpofüüsi ja käbinääret. Keskajus on nelja mugula kujul olev quadrigemina. Kaks eesmist on ühendatud visuaalne analüsaator, kaks tagumist - koos kuulmis.
Keskse evolutsiooni peamised etapid närvisüsteem kajastuvad ka inimese ontogeneesis. Neurulatsiooni staadiumis asetatakse närviplaat maha, muutudes sooneks ja seejärel toruks. Toru eesmine ots moodustab kõigepealt kolm aju vesiikulit: eesmine, keskmine ja tagumine. . Pärast seda jagatakse eesmine vesiikul kaheks, diferentseerudes eesajuks ja vaheajuks - keskmisest ajupõiekest areneb keskaju ning tagumisest vesiikulist tagaaju ja piklikaju.
Monogeenne pärimise tüüp. Näited.
Polügeensed või multifaktoriaalsed haigused. Pärimise tunnused.
Kahepaiksed (kahepaiksed).
Eesaju kahepaiksed moodustavad kaks poolkera, mille sees paiknevad külgvatsakesed koroidpõimikutega. Eesaju ees asuvad suured lõhnasibulad. Nad on poolkeradest nõrgalt piiritletud ja sabata kahepaiksetel kasvavad nad koos piki keskjoont. Haistmissibulatest tulevaid signaale analüüsitakse eesajus, mis on sisuliselt kõrgeim haistmiskeskus. Eesaju katuse moodustab esmane ajuvõlv - arhipallium. See sisaldab närvikiude ( valge aine) ja sügavuses nende all asuvad närvirakud. Eesaju põhjas on neuronite kobarad - striataalsed kehad.
Vahetult eesaju poolkerad asuvad taga vahepea hästiarenenud ülemise peaajulisandiga - käbinääre ja alumise ajulisandiga - ajuripatsiga. Üldjoontes sarnaneb vahelihas kalade omaga.
Kahepaiksete aju suurim osa on keskaju. See näeb välja nagu kaks koorega kaetud poolkera. See hõlmab nägemisnärvi jätkuna nägemistrakti ja siin on visuaalne analüsaator integreeritud teiste sensoorsed rajad ja moodustub keskus, mis täidab keerulisi assotsiatiivseid funktsioone. Seega on keskaju kesknärvisüsteemi juhtiv osakond, kus analüüsitakse saadud informatsiooni ja genereeritakse vastuseimpulsse, s.t. Kahepaiksetel, nagu kaladel, on ihtiopsiidi tüüpi aju.
Riis. 7. Konna aju (seljavaade).
1 – eesaju poolkerad.
2 – haistmissagarad.
3 – haistmisnärvid.
4 – vahepea.
5 – keskaju.
6 – väikeaju.
7 – piklik medulla.
8 – seljaaju.Väikeaju enamikul saba- ja sabata kahepaiksetel väikesed suurused ja sellel on pikliku medulla rombikujulise lohu esiservas põikhari. Väikeaju kehv areng peegeldab kahepaiksete motoorset koordinatsiooni. Suurem osa väikeajust koosneb keskmisest osast (väikeaju kehast), kuhu on integreeritud lihasretseptorite ja vestibulaarsüsteemi signaalid.
Kahepaiksetel, nagu kaladel, on väikeaju närvikiud seotud keskaju, ajutüve ja seljaajuga. Vestibulaar- ja väikeaju ühendused määravad loomade võimet keha liigutusi koordineerida.
Medulla põhitunnuste poolest sarnanevad kalade medulla piklikule. Seal on 10 paari kraniaalnärve, mis pärinevad ajutüvest.
Roomajad (roomajad).
Roomajad on tõelised maismaaloomad, kes võivad elada, paljuneda ja areneda veekogudest kaugel. Need kuuluvad kõrgemate selgroogsete hulka. Nende närvisüsteem on nende aktiivse ja keerulise eluviisi tõttu paremini arenenud kui kahepaiksetel.

Riis. 8. Sisaliku aju (Parkeri järgi).
A – pealtvaade.
B – altvaade.
B – külgvaade.
1 – eesaju; 2 – juttkeha; 3 – keskaju; 4 – väikeaju; 5 - medulla piklik; 6 – lehter; 7 – hüpofüüsi; 8 - chiasma; 9 – haistmissagarad; 10 – hüpofüüsi; II – XII- peanärvid.Eesaju mõõtmetelt oluliselt suurem kui kahepaiksed ja keerukama ehitusega; suureneb nende võime moodustada konditsioneeritud reflekse, kiiremini tekivad uued sidemed väliskeskkonnaga ning nad suudavad kohaneda keskkonnamuutustega paremini kui nende esivanemad. Eesaju koosneb kahest poolkerast, mis tahapoole kasvades katavad vahekeha, välja arvatud käbinääre ja parietaalelund. Eesaju laienemine toimub peamiselt striataalsete kehade (neuronite klastrite) tõttu, mis asuvad külgmiste vatsakeste põhjas. Nad toimivad kõrgema integratsioonikeskusena, analüüsides eesajusse sisenevat teavet ja arendades vastuseid. Seega lakkab see olemast ainult haistmiskeskus. Seda tüüpi aju nimetatakse sauropsid. Mis puutub ajuvõlvi, siis selles toimuvad olulised muutused. Esiaju katuse mõlemas poolkeras tekivad esimest korda evolutsioonis kaks halli aine saart (kortikaalsed rudimendid) – üks neist paikneb poolkerade mediaalsel ja teine lateraalsel küljel. Funktsionaalselt on oluline vaid mediaalne insula, mis esindab kõrgeimat haistmiskeskust. Üldiselt on ajukoore saared primitiivse ehitusega ja neid nimetatakse iidseks korteksiks (archicortex). Enamik autoreid peab ajukoore saari ühekihilisteks, kuigi krokodillidel võib eristada kahte või isegi kolme kihti.
Eesajuga seotud haistmisagarad on hästi arenenud. Mõnel liigil on nad istuv asend, kuid sagedamini eristatakse neid sibulateks ja varteks.
Roomajate eesaju uurimine on tehtud oluline evolutsioonilise neurohistoloogia jaoks, sest need on selgroogsete evolutsiooni võtmepunktiks, millest alates kulges eesaju areng kahes põhimõtteliselt erinevas suunas: mööda striataalset rada, kus lindudel domineerivad subkortikaalsed struktuurid, ja mööda kortikaalset rada, kus domineerivad ajukoore struktuurid. imetajatel.
Diencephalonõhukesel katusel on sellel kaks mullilaadset moodustist, millest üks asub ees ja seda nimetatakse parietaal- ehk parapineaalorganiks ning teine tagapool on epifüüs (käbinääre). Parapineaalelund täidab valgustundlikku funktsiooni ja seetõttu nimetatakse seda ka parietaalseks silmaks. Tegelikult moodustavad parapineaalelund ja käbinääre tandemi, mis on loomade igapäevase tegevuse regulaator. Parietaalset organit ei leidu aga kõigil roomajatel. Sellistel juhtudel on igapäevase tegevuse reguleerimise mehhanism erinev: teave pikkuse kohta päevavalgustund pärineb mitte parapineaalorganist, vaid visuaalne süsteem.
Keskaju mida esindab kolliikul ja oma põhitunnuste poolest on sellel sama korraldus, mis on omane kahepaiksetele, kuid roomajaid iseloomustab keskaju iga sensoorse süsteemi täpsem topograafiline esitus. Pealegi peaaegu kõike motoorsed tuumad väikeaju ja piklik medulla interakteeruvad keskaju katuse neuronitega. Samal ajal on keskaju kaotamas oma tähtsust kesknärvisüsteemi peamise integreeriva osakonnana. See funktsioon läheb üle eesajusse.
Mõned nägemis- ja kuulmiskiud saadetakse mööda möödaviiguteid, mööda keskajust, eesajusse. Keskajus saadakse keha automaatsete kaasasündinud reaktsioonide tagamise keskused ühtlaselt varajased staadiumid selgroogsete areng. Uued eesaju keskused võtavad üle teekumi funktsioonid ja moodustavad uusi motoorseid radu.
Väikeaju roomajate kõndimise ja jooksmise arenemise tõttu on see paremini arenenud kui kahepaiksetel. See koosneb kesksest aksiaalsest osast, mida nimetatakse ussiks, ja mõnel on külgmised labad. Väikeajule on iseloomulikud arvukad ühendused närvisüsteemi teiste liikumisega seotud osadega. Siin seotud spinotserebellaarne trakt, mis esineb nii kaladel kui ka kahepaiksetel, ulatub vermise seljaosani. Väikeaju ühendused on vestibulaarse aparatuuri, keskaju ja rhombencephaloniga. Vestibulaar- ja väikeaju ühendused kontrollivad keha asendit ruumis ja talamuse ühendused reguleerivad lihaste toonust.
Medulla moodustab järsu kurvi vertikaaltasapinnas, mis on üldiselt iseloomulik amnionitele.
Seal on 12 paari kraniaalnärve, mis pärinevad ajutüvest.
Linnud.
Linnud on kõrgelt spetsialiseerunud selgroogsete rühm, kes on kohanenud lennuks. Olles füsioloogiliselt roomajatele lähedased, on lindude aju paljuski sarnane roomajate omaga, kuigi nad erinevad mitmete progressiivsete tunnuste poolest, mis viisid keerukama ja mitmekesisema käitumiseni. Nende aju on üsna suur. Kui roomajatel on selle mass ligikaudu võrdne seljaaju massiga, siis lindudel on see alati suurem. Aju kõverad on teravalt väljendunud.
Aju laienemine on eelkõige tingitud arengust eesaju, mis katab sisuliselt kõik ajuosad, välja arvatud väikeaju. Tänu sellele ei ole keskaju ülalt näha, kuigi see on hästi arenenud. Eesaju katus on endiselt halvasti arenenud. Cora mitte ainult ei saa edasine areng, kuid poolkerades kaob ajukoore külgmine saar ja mediaalne säilitab kõrgema haistmiskeskuse tähtsuse.
Eesaju suuruse suurenemine toimub tänu põhjale, kus asuvad suured striataalsed kehad, mis on aju juhtiv osa. See tähendab, et linnud hoiavad sauropsid aju tüüp.
Haistmissagarad on väga väikesed, mis on seotud haistmismeele kehva arenguga, ja paiknevad tihedalt eesajuga.
Diencephalon väike. Enamikul lindudel säilib tavaliselt ainult käbinääre ja parapineaalelund kaob hilises embrüonaalses perioodis. Info päevavalguse pikkuse kohta ei tule parapineaalorganist, vaid otse nägemissüsteemist. Taalamuses on enim arenenud seljaosa, mis on aferentsete ühenduste lülituskeskus eesajuga. See sisaldab eesaju motoorsetest keskustest laskuvate kiudude teel tuumade kompleksi. Selle all on hüpotalamus, mis on ühendatud hüpofüüsiga, mis on lindudel hästi arenenud. Hüpotalamus mängib olulist rolli organismi hormonaalses regulatsioonis, säilitades homöostaasi, seksuaalset ja söömiskäitumist.
Keskaju koosneb kahest suurest küngast. See sisaldab kõrgeimaid keskusi visuaalse ja kuuldava teabe töötlemiseks, samuti liigispetsiifiliste pärilike koordineeritud reaktsioonide vormide reguleerimise keskusi, mis on elu aluseks. Samuti on sensoorsed tuumad, mis täidavad assotsiatiivset funktsiooni, saates signaale vahe- ja eesajusse. Samal ajal väheneb laskuvate ühenduste arv keskaju katuse ja motoorsete keskuste vahel, kuna lindudel tekivad vööt-retikulaarsed suhteliselt autonoomsed ühendused eesaju striataalkehade ja eesaju kehade vahel. retikulaarne moodustumine ajutüvi.
Väikeaju suur ja selle struktuur muutub keerulisemaks. Ees puutub see kokku eesaju poolkerade tagumiste servadega ja taga katab olulise osa piklikest medullast. Väikeaju jaguneb keskmiseks osaks (vermis) ja külgmisteks projektsioonideks. Selle uss on täpiline iseloomulike põikisuunaliste soontega. Väikeaju suhteliselt keeruline struktuur on tingitud keerulistest liigutustest, mis nõuavad lennu ajal kõrget koordinatsiooni.Medulla suhteliselt väike, selle alumine külg, isegi rohkem kui roomajatel, moodustab allapoole kõveruse ja vahekeha piirkonnas on ülespoole painutamine.
Lindude kraniaalnärvid on esindatud 12 paarina.
Imetajad.
Imetajad on kõrgelt arenenud kesknärvisüsteemiga kõige paremini organiseeritud selgroogsete klass. Sellega seoses on imetajate kohanemisreaktsioonid keskkonnatingimustega keerulised ja väga arenenud.
Eesaju suur, on see oluliselt suurem kui kõik teised ajuosad. Selle poolkerad kasvavad igas suunas, varjates vahepead. Aju keskaju on väljastpoolt nähtav ainult platsentadel ja madalamatel platsentadel ning kabiloomadel, lihasööjatel, vaalalistel ja primaatidel on see kaetud tagumise osaga. ajupoolkerad. Inimloomadel ja inimestel kuklasagarad eesaju surutakse väikeajule.
Kui algselt, evolutsiooni käigus, moodustasid telentsefaloni põhiosa haistmissagaratest, siis imetajatel on haistmissagarad välja arenenud vaid madalamatel ning kõrgematel on haistmisagar väikeste lisandite kujul, mis jagunevad haistmisagaraks ja haistmistrakt.
Imetajate eesaju suhtelise suuruse suurenemine on seotud peamiselt selle katuse, mitte aga juttkeha kasvuga, nagu lindudel. Medullaarne võlv (katus) on moodustatud hallainest, mida nimetatakse ajukooreks. Viimane on kompleks, mis koosneb iidsest kuubist (paleopalium), vanast kuubist (archipallium) ja uuest kuubist (neopalium). Uus kuub on vahepealsel positsioonil, paiknedes vana ja iidse kuube vahel. Vana vahevöö ehk vana ajukoor paikneb mediaalselt ja vanasti nimetati seda hipokampuseks ehk Ammoni sarveks. Iidne kuub ehk iidne koor on külgmisel positsioonil.
Uut mantlit nimetatakse tavaliselt neokorteksiks ( neokorteks) ja just sellest koosnevad peamiselt eesaju poolkerad. Sel juhul võib poolkerade pind olla sile (lisentsefaalne) või volditud (soonte ja keerdudega). Lisaks sellele eristatakse poolkerades sellest hoolimata 4 kuni 5 laba. Eesaju labadeks jagamise põhimõte põhineb teatud soonte ja keerdude topograafial. Jaotumine labadeks lisefaalses (sileda) ajus on tingimuslik. Tavaliselt on parietaalsagarad, temporaal-, kuklaluu- ja otsmikusagarad ning kõrgematel primaatidel ja inimestel on ka viies sagar, mida nimetatakse insulaks. See moodustub embrüonaalsel perioodil tänu oimusagara kasvule poolkerade ventraalsele küljele.Võttes ajupoolkerade esialgseks tüübiks lisefaalse aju, eristatakse soonte mustri arendamiseks kolme võimalust: pikisuunaline, kaarjas ja “primaadi tüüp”. Primaadi tüübi puhul on otsmikusagara soon suunatud rostraalselt ja oimusagarates ventro-dorsaalselt
Aju kuju võib oluliselt mõjutada sulci ja gyri asukohta. Enamikul imetajatel on aju piklik rostro-saba suunas. Kuid paljudel delfiinidel on aju külgsuunas laienenud ja suhteliselt lühenenud.
Iseloomustamaks imetaja eesaju suur tähtsus, lisaks vagudele ja keerdudele on jaotusmuster neuronaalses ajukoores (tsütoarhitektuur). Imetajate neokorteksil on kuuekihiline struktuur ja seda iseloomustab püramiidrakkude olemasolu, mis teiste selgroogsete ajus puuduvad. Eriti suuri püramiidrakke (Betzi rakke) leidub motoorses ajukoores. Nende aksonid edastavad närviimpulsse seljaaju motoorsetele neuronitele ja kraniaalnärvide motoorsete tuumade motoorsetele neuronitele.
Ajukoore erinevad piirkonnad on eripiirkonnad erinevatest meeleorganitest tuleva teabe töötlemiseks. Seal on sensoorsed ja motoorsed alad. Viimased moodustavad närvikiudude laskumisteed ajutüve ja seljaaju motoorsete tuumadeni. Ajukoore tundliku ja motoorsete piirkondade vahel on integratiivsed piirkonnad, mis ühendavad ajukoore sensoorse ja motoorsete piirkondade sisendid ning määravad kindlaks spetsiifiliste liigispetsiifiliste funktsioonide toimimise. Lisaks on ajukoores assotsiatiivseid tsoone, mis ei ole seotud konkreetsete analüsaatoritega. Need kujutavad endast pealisehitist ajukoore ülejäänud piirkondades, pakkudes mõtteprotsesse ja salvestades spetsiifilist ja individuaalset mälu.
Kogu ajukoores jaotatud tsoonide kompleks on seotud väljade funktsionaalse spetsialiseerumisega. Sel juhul langevad väljade morfoloogilised ja funktsionaalsed piirid üsna täpselt kokku. Konkreetse välja tuvastamise kriteeriumiks on rakuliste elementide jaotumise muutus ajukoores või uue alamkihi tekkimine selles.
Teatud valdkondade arhitektoonika tunnused on nende funktsionaalse spetsialiseerumise morfoloogiline väljendus. Põldude tsütoarhitektoonika muutuse põhjuseks on tõusvate ja laskuvate närvikiudude arvu suurenemine. Nüüdseks on inimeste ja paljude laboriloomade jaoks loodud põldude topoloogilised kaardid.
Ajukoore väljad on osa teatud sagaratest ja on ise jagatud funktsionaalseteks tsoonideks, mis on seotud konkreetsete elundite või nende osadega ja millel on korrastatud sisemine struktuur. Igal põllul või tsoonis on nö moodulid ajukoore organisatsiooni vertikaalne korrastatus. Moodul on kas kolonni või glomeruli kujul, mis sisaldab kogu ajukoore paksuses paiknevaid neuroneid. Kolonn sisaldab 110 neuroni rühma, mis paiknevad kapillaaride paari vahel, mis kulgevad üle ajukoore läbimõõdu.
Kõige iidsemate hominiidide aju moodustumise staadiumis oli piirkond, kuhu loodusliku valiku tegevus oli suunatud, ajukoor ja ennekõike selle järgmised sektsioonid: alumine parietaalne, alumine eesmine ja temporo-parietaalne piirkond. Ellujäämise eelised said need indiviidid ja seejärel need esilekerkivate inimeste populatsioonid, kes osutusid ajukoore osade mõne elemendi arengus edasijõudnuks ( suurem ala mitmekesisemate ja mobiilsemate ühendustega väljad, paranenud vereringe tingimused jne). uute ühenduste ja struktuuride arendamine ajukoores andis uusi võimalusi tööriistade valmistamiseks ja meeskonna moodustamiseks. Aju arengule aitasid omakorda kaasa tehnoloogia uus tase, kultuuri ja kunsti algus loodusliku valiku kaudu.
Tänaseks on kujunenud ettekujutus inimese eesajukoore spetsiifilisest süsteemsest kompleksist, mis hõlmab ajukoore alumisi parietaalseid, tagumisi ülemisi temporaalseid ja alumisi otsmikusagaraid. Seda kompleksi seostatakse kõrgemate funktsioonidega – kõne, tööaktiivsus ja abstraktne mõtlemine. Üldiselt on see teise signaalisüsteemi morfoloogiline substraat. Sellel süsteemil ei ole oma perifeerseid retseptoreid, vaid kasutatakse erinevate meeleorganite vana retseptoraparaati. Näiteks on kindlaks tehtud, et keelel on kompimisaparaadi spetsiaalne osa, mille areng määrab lapse artikuleeritud kõne kujunemise algfaasis heli tekitamise järjestuse.
Eesaju submantlistruktuuride hulka kuuluvad basaalganglionid, juttkeha (iidne, vana ja uus) ja vaheseinaväli.
Eesaju ja vaheaju erinevates osades on morfofunktsionaalsete struktuuride kompleks nn Limbiline süsteem. Viimasel on arvukalt seoseid neokorteksi ja autonoomse närvisüsteemiga. See ühendab aju funktsioone, nagu emotsioonid ja mälu. Limbilise süsteemi osa eemaldamine toob kaasa looma emotsionaalse passiivsuse ja selle stimuleerimine hüperaktiivsuseni. Kõige olulisem funktsioon Limbiline süsteem suhtleb mälumehhanismidega. Lühiajaline mälu on seotud hipokampusega ja pikaajaline mälu neokorteksiga. Limbilise süsteemi kaudu ammutatakse neokorteksist looma individuaalne kogemus, toimub siseorganite motoorne kontroll ja looma hormonaalne stimulatsioon. Veelgi enam, mida madalam on neokorteksi arengutase, seda rohkem sõltub looma käitumine limbilisest süsteemist, mis viib emotsionaalse-hormonaalse kontrolli domineerimiseni otsuste tegemise üle.
Imetajatel võimaldavad neokorteksist limbilise süsteemi laskuvad ühendused integreerida mitmesuguseid sensoorseid sisendeid.
Roomajate ajukoore esimeste algte ilmumisega eraldus vahevöö kommissuurist väike närvikiudude kimp, mis ühendas vasakut ja paremat poolkera. Platsenta imetajatel on selline kiudude kimp palju arenenum ja seda nimetatakse corpus callosumiks. Viimane tagab poolkeradevahelise side funktsiooni.
Diencephalon, nagu ka teised selgroogsed, koosneb epitalamusest, talamusest ja hüpotaalamust.
Neokorteksi areng imetajatel tõi kaasa talamuse ja ennekõike seljaosa järsu suurenemise. Taalamuses on umbes 40 tuuma, milles tõusuteed lülituvad viimastele neuronitele, mille aksonid jõuavad ajukooresse, kus töödeldakse kõikidest sensoorsetest süsteemidest tulevat infot. Samal ajal töötlevad eesmised ja külgmised tuumad ning edastavad vastavatele visuaalsete, kuulmis-, puute-, maitse- ja interotseptiivsete signaalide. projektsioonitsoonid koor. On olemas arvamus, et valutundlikkust ei projitseerita eesaju poolkerade ajukooresse, vaid selle kesksed mehhanismid asuvad talamuses. See oletus põhineb asjaolul, et ajukoore erinevate piirkondade ärritus ei põhjusta valu, talamuse ärritus aga valu. tugev valu. Osa talamuse tuumadest lülituvad ja teine osa on assotsiatiivsed (neist viivad teed ajukoore assotsiatiivsetesse tsoonidesse). Taalamuse mediaalses osas on tuumad, mis madala sagedusega elektrilise stimulatsiooniga põhjustavad ajukoores inhibeerivate protsesside arengut, mis viib une. Nende tuumade kõrgsageduslik stimulatsioon põhjustab kortikaalsete mehhanismide osalist aktiveerimist. Seega osaleb talamokortikaalne reguleerimissüsteem, kontrollides tõusvate impulsside voogu, une ja ärkveloleku vahelise muutuse korraldamises.
Kui madalamatel selgroogsetel asuvad kõrgemad sensoorsed ja assotsiatsioonikeskused keskajus ning dorsaalne talamus on tagasihoidlik integraator keskaju ja keskaju vahel. haistmissüsteem, siis on see imetajatel kõige olulisem kuulmis- ja somatosensoorsete signaalide ümberlülitamise keskus. Samal ajal on somatosensoorsest piirkonnast saanud vahekeha kõige silmapaistvam moodustis ja see mängib liigutuste koordineerimisel tohutut rolli.
Tuleb märkida, et talamuse tuumade kompleks moodustub nii vaheaju primordiumi tõttu kui ka migratsiooni tõttu keskajust.
Hüpotalamuse moodustab arenenud külgmised eendid ja õõnes vars - lehter. Viimane lõpeb tagumises suunas, kusjuures neurohüpofüüs on tihedalt ühendatud adenohüpofüüsiga.
Hüpotalamus on kõrgeim reguleerimiskeskus endokriinsed funktsioonid keha. See ühendab endokriinsed regulatsioonimehhanismid närvisüsteemiga. Lisaks on see autonoomse närvisüsteemi sümpaatilise ja parasümpaatilise osakonna kõrgeim keskus.
Epitalamus toimib igapäevase ja hooajalise aktiivsuse neurohumoraalse regulaatorina, mis on kombineeritud loomade puberteedi kontrollimisega.
Keskaju moodustab quadrigeminaalse piirkonna, mille eesmised tuberklid on ühendatud visuaalse analüsaatoriga ja tagumised kuulmisosaga. Eesmise ja tagumise mugulate suhtelise suuruse suhte järgi saab otsustada, milline süsteem, kuulmis- või visuaalne, on ülekaalus. Kui eesmised tuberkulid on paremini arenenud, tähendab see visuaalset aferentatsiooni (kabiloomad, paljud röövloomad ja primaadid), kui tagumised, siis kuulmisaferentatsiooni (delfiinid, nahkhiired jne).
Märgistus jaguneb sensoorseks ja motoorseks tsooniks. Motoorses tsoonis on kraniaalnärvide motoorsed tuumad ning laskuvad ja tõusvad spinotserebraalsed kiud.
Seoses neokorteksi arenguga imetajatel kõrgema integratsioonikeskusena, võimaldasid keskaju kaasasündinud reaktsioonid ajukoorel "mitte haakuda". primitiivsed vormid liigispetsiifilised reaktsioonid välistele signaalidele, samas kui keerukad assotsiatiivsed funktsioonid on omandatud spetsiaalsete kortikaalsete väljade poolt.
Väikeaju imetajatel omandab see kõige keerulisema struktuuri. Anatoomiliselt võib selle jagada keskmiseks osaks - vermis, selle mõlemal küljel paiknevad poolkerad ja flokulonodulaarsed labad. Viimased esindavad fülogeneetiliselt iidset osa – Archicerebellum. Poolkerad jagunevad omakorda ees- ja tagasagarateks. Poolkerade eesmised labad ja väikeaju vermise tagumine osa esindavad fülogeneetiliselt vana väikeaju – paleokerebellumi. Fülogeneetiliselt hõlmab väikeaju noorim osa, neotserebellum, väikeaju poolkerade tagumiste labade esiosa.
Väikeaju poolkerad jagunevad ülemiseks pinnaks, mis moodustab väikeaju ajukoore, ja närvirakkude klastriteks - väikeaju tuumadeks. Väikeaju ajukoor on ehitatud ühe põhimõtte järgi ja koosneb 3 kihist. Väikeaju on kesknärvisüsteemi teiste osadega ühendatud kolme paari varrega, mis on moodustatud närvikiudude kimpudest. Tagajalad koosnevad peamiselt propriotseptiivsetest kiududest, mis pärinevad seljaajust. Keskmised varred koosnevad väikeaju ja eesaju koort ühendavatest kiududest ning eesmised varred moodustuvad väikeaju ja keskaju ühendavatest laskuvatest kiududest.Vestibulotserebellaarsed ühendused määravad loomade võime koordineerida keha liigutusi, mis on väikeaju põhifunktsioon. Lisaks on väikeaju hambatuuma tekkimise tõttu imetajatel tekkinud uued võimsamad väikeaju teed. See võtab vastu kiudaineid väikeaju poolkerade erinevatest osadest ja edastab signaale talamusele, kus sensomotoorsed signaalid on integreeritud eesaju ajukoore keskuste aktiivsusega
... funktsionaalne fülogeneesia morpho-funktsionaalne... keskne närviline, endokriinsed, hingamisteede ja teised süsteemidFunktsionaalne olek...
Alates fundamentaalteadustest morfoloogiast, füsioloogiast, füüsikast, evolutsiooniõpetusest – intelligentsete süsteemide ökoloogia, meditsiini ja rahvaterviseni (1)
Raamat... funktsionaalne, parameetrilised, sünergilised omadused fülogeneesia ja inimese aju ontogenees. Teadmiste kogum selle kohta morpho-funktsionaalne... keskne närviline, endokriinsed, hingamisteede ja teised süsteemidFunktsionaalne olek...
Alates fundamentaalteadustest morfoloogiast, füsioloogiast, füüsikast, evolutsiooniõpetusest – intelligentsete süsteemide ökoloogia, meditsiini ja rahvaterviseni (2)
Raamat... funktsionaalne, parameetrilised, sünergilised omadused fülogeneesia ja inimese aju ontogenees. Teadmiste kogum selle kohta morpho-funktsionaalne... keskne närviline, endokriinsed, hingamisteede ja teised süsteemidFunktsionaalne olek...
Kesknärvisüsteemi füsioloogia (1)
DokumentKodeerimine - muutumine teave sisse... selgrootud ja madalam selgroogsed loomad... ühendus närviline Ja endokriinsedsüsteemid. ... funktsionaalne süsteemid, mis võtavad vastu signaale elundidtundeid pärast eeltöötlust keskses närviline ...
Boriss Gurjevitš Meštšerjakov Vladimir Petrovitš Zinchenko Suur psühholoogiline sõnastik Sisu
Dokument... elundidtundeid(Ja Inimkehaüldiselt) kombinatsioonis funktsionaalne asümmeetria... närviline rakud ( närviline G.) väljaspool c. n. Koos. U selgroogsed loomad G. paiknevad piki seljaaju ja sisemiste seintes elundid. U selgrootud ...


