Razvoj mozga vodozemaca. Klasa vodozemci ili vodozemci
Vodozemci ili vodozemci, kao odrasli, obično su kopnene životinje, ali su i dalje usko povezane s vodenim okolišem, a njihove ličinke stalno žive u vodi. Shodno tome, ruski i grčki (vodozemci - vodeći dvostruki život) imena odražavaju glavne karakteristike ovih kičmenjaka. Vodozemci potječu, kao što je već spomenuto, od devonskih riba s režnjevima peraja koje su živjele u malim slatkovodnim tijelima i puzale do obale uz pomoć svojih mesnatih uparenih peraja.
Vanjski objekat. Tijelo (sl. 147) se sastoji od glave, trupa, prednjih i stražnjih parnih raskomadanih udova. Udovi se sastoje od tri dijela: prednji - od ramena, podlaktice i šake, stražnji - od butine, potkolenice i stopala. Samo manjina modernih vodozemaca ima rep (red repa - tritoni, daždevnjaci, itd.). Smanjuje se kod odraslih oblika najveće grupe vodozemaca - anurana (žabe, žabe i dr.) zbog prilagodbe potonjih na kretanje skakanjem po kopnu, ali je očuvan u njihovim ličinkama - punoglavcima koji žive u vodi. Kod nekoliko vrsta koje vode polu-podzemni način života (red bez nogu ili cecilijani), udovi i rep su smanjeni.
Glava je pokretno zglobljena s tijelom, iako je njen pokret vrlo ograničen i nema izraženog vrata. Karakteristični su raskomadani udovi i pokretna veza između glave i tijela kopnenih kralježnjaka, oni su odsutni u ribama. Tijelo kopnenih formi je spljošteno u dorzo-ventralnom smjeru, dok je kod riba (zbog njihove adaptacije na plivanje) u pravilu stisnuto bočno. Kod vodenih vodozemaca oblik tijela je sličan ribljem. Veličina tijela se kreće od 2 do 160 cm (japanski daždevnjak); u prosjeku, vodozemci su manje veličine od ostalih kopnenih životinja. Koža gola bogata žlezdama, odvojeno na mnogim mjestima od mišića zbog prisustva potkožnih limfnih šupljina. Opremljen je velikim brojem krvni sudovi a također obavlja i respiratornu funkciju (vidi dolje). Kod nekih vrsta pražnjenja kožne žlezde otrovno. Boja kože je veoma raznolika.
Nervni sistem. U vezi s prilagodbom vodozemaca životu na kopnu, a posebno u vezi s radikalnom promjenom prirode kretanja, nervni sistem se dosta promijenio. Prednji mozak kod vodozemaca (vidi sliku 133, B) je veći od prosjeka; kod riba se u pravilu opaža suprotan omjer. To se objašnjava činjenicom da ribe imaju funkcije prednji mozak povezani su samo s percepcijom olfaktornih podražaja kod vodozemaca, počinje sudjelovati u koordinaciji različitih tjelesnih funkcija, a u njegovom površinskom sloju pojavljuju se rudimenti korteksa (još uvijek vrlo slabi) u kojima su koncentrirane živčane stanice. Istovremeno, treba napomenuti da su olfaktorni režnjevi dobro razvijeni u prednjem mozgu. Mali mozak kod vodozemaca je vrlo slabo razvijen, za razliku od riba. Ribe se stalno kreću, a njihov položaj tijela je nestabilan, dok su vodozemci, oslonjeni na noge, u prilično stabilnom položaju. Parcele kičmena moždina, na mjestima gdje od njega odlaze živci, idući do mišića nogu, koji obavljaju mnogo više posla od mišića parnih peraja riba, zadebljani su i na njih su povezani brahijalni i lumbalni pleksusi živaca. Periferni nervni sistem se uvelike promijenio zbog diferencijacije mišića (vidi dolje) i pojave dugih, spojenih udova.

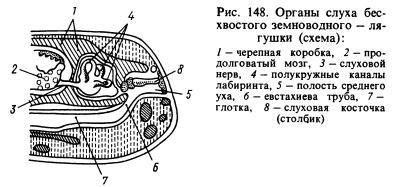
Od čulnih organa, organ sluha je pretrpio najznačajnije promjene. Prijenos zvučnih valova iz vodene sredine u životinjska tkiva, koja su također zasićena vodom i imaju približno ista akustička svojstva kao voda, odvija se mnogo bolje nego iz zraka. Zvučni talasi koji putuju vazdušno okruženje, gotovo u potpunosti se reflektiraju od površine životinje i samo oko 1% energije ovih valova prodire u njeno tijelo. U tom smislu, vodozemci su se razvili pored lavirinta, ili unutrasnje uho, novi dio slušnog organa je srednje uho. To je (Sl. 148) mala šupljina ispunjena zrakom, koja komunicira sa usnom šupljinom kroz Eustahijevu cijev i zatvorena je izvana tankom, elastičnom bubnom opnom. U srednjem uhu nalazi se slušna ploča (ili stub) koja se jednim krajem naslanja na bubnu opnu, a drugim na prozor prekriven filmom i vodi u lobanjsku šupljinu, gdje se nalazi labirint okružen perilimfom. Pritisak unutar srednjeg uha jednak je atmosferskom pritisku i bubna opna može vibrirati pod uticajem zvučnih vazdušnih talasa, čiji se uticaj dalje prenosi preko slušne koščice i perilimfe na zidove lavirinta i opaža se na završecima slušnog živca. Šupljina srednjeg uha formirana je od prvog škržnog proreza, a stub je formiran od hiomandibularne kosti (hiomandibularne kosti) smještene u blizini proreza, koja je visceralni dio lubanje objesila na mozak gdje se nalazio labirint iza ušne kosti.
Oči su prekrivene pokretnim kapcima, koji štite organe vida od isušivanja i začepljenja. Zahvaljujući promjenama u obliku rožnice i sočiva, vodozemci vide dalje od riba. Vodozemci dobro percipiraju male promjene temperature. Osetljivi su na uticaje razne supstance, rastvoren u vodi. Njihov njušni organ reagira na iritacije uzrokovane plinovitim tvarima. Tako su osjetilni organi vodozemaca pretrpjeli niz promjena u vezi s prelaskom na život na kopnu. Ličinke i odrasle životinje koje stalno žive u vodi imaju, kao i ribe, organe bočne linije.
Vodozemce karakteriziraju prilično složene instinktivne radnje, posebno tokom sezone parenja. Na primjer, muška babica krastača, koja živi u Rusiji u zapadnoj Ukrajini, omota "vezice" jaja oko svojih stražnjih udova i skriva se na osamljenim mjestima na obali dok se punoglavci ne razviju. Nakon 17-18 dana mužjak se vraća u vodu, gdje se izlegu punoglavci. Ovo je svojevrsni instinkt za zaštitu potomstva. Još složeniji instinkti poznati su kod brojnih tropskih vodozemaca bez repa. Vodozemci također imaju uslovne reflekse, ali se oni razvijaju s velikim poteškoćama.
Motorni sistem i skelet. Mišićni sistem u vezi sa razni uređajiživot na kopnu (razvoj udova kopnenog tipa, pojava pokretnog zgloba između glave i tijela, itd.) doživio je radikalne transformacije, iako je zadržao mnoge osobine svojstvene ribama. Mišićni sistem riba je vrlo ujednačen i uglavnom se sastoji od sličnih bočnih mišićnih segmenata. Kod vodozemaca, mišićni sistem je postao više diferenciran i sastoji se od raznih mišića (Sl. 149). Vodozemci su postavili temelje mišićnog sistema, koji se kasnije razvio i složeniji kod pravih kopnenih kičmenjaka - gmizavaca, ptica i sisara. Ovo se odnosi i na skelet.
Lobanja vodozemaca ima mnogo hrskavičnih elemenata, što se vjerovatno objašnjava potrebom za smanjenjem tjelesne težine zbog poluzemaljskog načina života. Lobanja sadrži mnogo kostiju navedenih u opisu lubanje viših riba, uključujući parasfenoid karakterističan samo za ribe i vodozemce (Sl. 150). Kako je hiomandibularna kost postala slušna kost, ulogu privjeska ima četvrtasta kost. Zbog gubitka škržnog aparata u odrasloj dobi, škržni lukovi su smanjeni i sačuvani su samo njihovi modificirani ostaci. Hioidni luk se jako mijenja i djelimično reducira. Lobanja vodozemaca je vrlo široka, što je dijelom posljedica karakteristika njihovog disanja. Donja vilica, kao i kod koščatih riba, sastoji se od nekoliko kostiju.
Kičmeni stub (Sl. 150) kod bezrepih životinja je vrlo kratak i završava se dugom kosti - urostilom, formiranom od rudimenata kaudalnih kralježaka. Kod repatih vodozemaca kaudalni dio kičmenog stuba sastoji se od niza pršljenova. Kod ovih vodozemaca rep igra značajnu ulogu u kretanju: u vodi se koristi za plivanje, na kopnu se koristi za održavanje ravnoteže. Rebra su slabo razvijena (kod repastih vodozemaca) ili su smanjena, a njihovi ostaci su srasli s poprečnim nastavcima kralježaka (kod drugih drevnih vodozemaca su imali rebra). Njihovo smanjenje u modernim oblicima objašnjava se potrebom da se smanji tjelesna težina (koja se znatno povećala prilikom prelaska iz vodene sredine u zrak) kod ovih kičmenjaka, koji još nisu dovoljno prilagođeni kretanju na kopnu. Zbog smanjenja rebara, vodozemci nemaju prsa. Prvi pršljen je drugačije strukturiran nego kod riba: ima dva glenoidna jama za artikulaciju sa dva okcipitalna kondila lubanje, zbog čega je glava vodozemaca postala pokretljiva.
Skelet prednjeg uda (Sl. 150) se sastoji od humerus, dvije kosti podlaktice - radijus i ulna, kosti ručnog zgloba, metakarpalne kosti i falange prstiju. Skelet zadnjeg ekstremiteta (Sl. 150) sastoji se od butine, dvije kosti potkolenice - tibije i fibule, tarzalnih kostiju, metatarzalnih kostiju i falangi prstiju. Posljedično, sličnost u građi oba para udova, unatoč određenim razlikama u njihovim funkcijama, vrlo je velika. U početku, prednje i zadnje noge su bile petoprsti moderni vodozemci mogu imati manje prstiju. Za plivanje se koriste i stražnji udovi mnogih vodozemaca bez repa, pa su stoga izduženi, a prsti su povezani plivačkim membranama.
Pojasevi udova su mnogo bolje razvijeni od onih kod riba. Rameni pojas se sastoji od koštanih i hrskavičnih elemenata: lopatica, ključna kost, vrana kost (korakoid) itd. (Sl. 150). Ključne kosti i korakoidi su povezani sa sternumom, koji takođe uključuje koštane i hrskavične elemente. Glava humerusa se artikulira sa ramenim pojasom. Stražnji pojas udova, ili karlica, sastoji se od tri kosti: iliuma, pubisa i išiuma (slika 150). Veliki acetabulum koji formiraju ove kosti služi za artikulaciju sa glavom femura. Zdjelica je povezana s jednim kralješkom - sakralnim, zahvaljujući kojem su zadnje noge, za razliku od trbušnih peraja ribe, dobile prilično jaku potporu.
Cirkulatorni sistem. Kod ličinki vodozemaca koji žive u vodi i dišu škrgama, krvožilni sistem je u osnovi sličan krvožilnom sistemu riba, ali se kod odraslih životinja koje vode kopneni način života značajno mijenja zbog zamjene škržnog disanja plućnim disanjem, pojačano disanje kože, te razvoj udova kopnenih životinja i druge promjene tijela. Srce (vidi sliku 134, B, 151) se sastoji od tri komore: desne i lijeve pretkomore i jedne komore. Polazi s desne strane potonjeg conus arteriosus(bio je prisutan i u ribama, precima vodozemaca), od kojih potiču četiri para arterija: prvi par - karotidnih arterija , nosioci krvi u glavu, drugi i treći par su žile koje se spajaju i formiraju najveći sud u tijelu - aorta, čije su grane usmjerene na različite dijelove tijela, četvrti par - plućne arterije, koje se zatim dijele na nezavisne kožne i plućne arterije.
Iz pluća oksigenirana krv teče kroz plućne vene u lijevu pretkomoru, a zasićena krv u sve dijelove tijela ugljen-dioksid, u prednjem delu tela uliva se u prednju šuplju venu, a u zadnjem delu tela u zadnju šuplju venu (Sl. 152). Obe šuplje vene se prazne u venski sinus, odakle ulazi krv (zasićena ugljičnim dioksidom). desna pretkomora. Iz oba atrija krv ulazi u jednu komoru srca. Unutrašnja površina ventrikula ima udubljenja i stoga krv u njoj nema vremena da se potpuno miješa: u lijevom dijelu je krv zasićena kisikom, u desnom dijelu je krv zasićena ugljičnim dioksidom, au srednjem dijelu miješano je. Budući da conus arteriosus počinje sa desna strana ventrikula, tada će prvi dio krvi koji ulazi u nju (tj. arterijski konus) biti venski, šalje se u najzadnje arterije - plućne.
Izmiješana krv koja zatim teče u arterije koje formiraju aortu, a kroz grane potonje u sve dijelove tijela. Oksigenirana krv iz lijeve strane komore šalje se u karotidne arterije. Ovome moramo dodati da krv, zasićena kisikom u koži, ulazi, kao što je gore navedeno, kroz prednju šuplju venu i venski sinus u desnu pretkomoru i tako razrjeđuje krv koja se tamo nalazi. venska krv, zatim gurnuti u sudove koji formiraju aortu. Posljedično, zahvaljujući gore opisanim uređajima, kao i ostalima koji ovdje nisu opisani, različiti dijelovi tijela primaju krv nejednako zasićenu kisikom. Na sl. 152 prikazuje glavnu arteriju i venske žile vodozemci.
Kod vodozemaca, zbog snažnog razvoja udova i veće disekcije tijela nego kod riba, mreža krvnih žila se značajno promijenila. Pojavilo se mnogo novih plovila kojih nije bilo u ribama, a pojavio se i sistem plovila karakterističnih za kopnene kralježnjake. Istovremeno, treba imati na umu da je cirkulacijski sustav vodozemaca mnogo jednostavniji od sustava viših kralježnjaka.
Respiratornog sistema. Skoro svi vodozemci imaju pluća (vidi sliku 151; 153). Ovi organi još uvijek imaju vrlo jednostavnu strukturu i predstavljaju vrećice tankih stijenki u čijim se zidovima grana prilično gusta mreža krvnih žila. Jer unutrašnji zid Pluća su gotovo glatka, tada je njihova površina relativno mala. Traheja je skoro nerazvijena, a pluća su direktno povezana sa larinksom. Budući da vodozemci nemaju grudni koš (vidi gore), čin disanja osigurava se radom mišića usne šupljine. Udisanje se dešava na sledeći način. Sa otvorenim nozdrvama (koje su, za razliku od nozdrva kod riba, prolazne, tj. pored vanjskih nozdrva postoje i unutrašnje nozdrve - choanae) I zatvorena usta Dno velike usne duplje se povlači unazad i u nju ulazi vazduh. Zatim se nosnice zatvaraju posebnim ventilima, dno usta se podiže i zrak se ubacuje u pluća. Izdisaj se javlja kao rezultat kontrakcije trbušnih mišića.
Vodozemci primaju značajnu količinu kisika kroz kožu i sluznicu usne šupljine. Neke vrste daždevnjaka uopće nemaju pluća i sva izmjena plinova se odvija kroz kožu. Međutim, koža može obavljati funkcije disanja samo ako je vlažna. Zbog toga je nemoguće da vodozemci žive u uslovima u kojima je vlažnost vazduha nedovoljna za njih. Larve koje žive u vodi dišu kroz škrge (prvo vanjske, a zatim unutarnje) i kožu. Neki repasti vodozemci koji stalno žive u vodi imaju škrge koje ostaju do kraja života. Dakle, u pogledu metoda disanja, vodozemci su još uvijek bliski ribama.
Ekskretorni sistem. Bubrezi (vidi sliku 136, A, B; sl. 151), kao i kod riba, su trup. Vukovi kanali se ulijevaju u kloaku. Tu se otvara mjehur, gdje se nakuplja urin. Uklanjanje produkata disimilacije također se događa kroz kožu i pluća.
Probavni sustav. Usna šupljina je veoma široka. Određene vrste (uglavnom repasti vodozemci) imaju mnogo malih, ujednačenih, primitivno raspoređenih zuba koji sjede na čeljustima, sošnicama, nepcima i drugim kostima i služe samo za držanje plijena. Kod većine vrsta (uglavnom vodozemaca bez repa) zubi su djelomično ili potpuno reducirani, ali im se jezik jako razvija. Potonji kod žaba je pričvršćen na prednjem kraju i može se baciti daleko naprijed sa zadnjim krajem kako bi se uhvatio plijen. Vrlo je ljepljiv i dobro prilagođen za obavljanje ove funkcije. Kod vrsta koje stalno žive u vodi, jezik je obično smanjen. Hvatanje plijena kod takvih vodozemaca vrši se čeljustima.
Digestivna cijev (vidi sliku 151) je relativno kratka i sastoji se od ždrijela, jednjaka, želuca, tankog crijeva i vrlo malog rektuma (debelog crijeva). Stražnji dio rektuma je kloaka; kroz njega se, osim izmeta, izlučuju urin i seksualni proizvodi. Žlijezde slinovnice, kojih nema u ribama, otiču se u usnu šupljinu. Sekret ovih žlijezda služi uglavnom za vlaženje hrane. Žlijezde pljuvačne žlijezde su vrlo slabo razvijene kod vrsta koje žive u vodi, a mnogo bolje kod kopnenih. Jetra je velika; pankreas je dobro definisan. Hrana odraslih vodozemaca je uglavnom životinjska (insekti, mali kralježnjaci itd.). Punoglavci vodozemaca bez repa uglavnom su biljojedi.

Reprodukcija. Muške gonade (testisi) leže blizu bubrega (vidi sliku 151, B). Njihovi kanali se otvaraju u tubule prednjeg dijela bubrega (vidi sliku 136, A) i sjeme se izlučuje, poput mokraće, kroz Wolfove kanale. Ženske spolne žlijezde (jajnici) uvelike rastu tokom perioda mrijesta. Jaja izlaze kroz veoma duge Müllerove kanale (vidi sliku 136, B). Potonji nemaju direktnu vezu sa jajnicima i jajne ćelije koje sazrijevaju kroz tjelesnu šupljinu ulaze u lijeve Müllerovih kanala.
Oplodnja se u većini slučajeva dešava u vodi. Kod mnogih vodozemaca tome prethodi zbližavanje mužjaka i ženke: mužjak stisne ženku s leđa, pritisne prednje udove na nju trbušni zid a to olakšava puštanje jaja u vodu, koje on odmah oplodi. Dakle, u prisustvu spolnog odnosa, do oplodnje dolazi izvan ženskog tijela. Kod manjine vrsta (na primjer, tritone), mužjak oslobađa sjeme u posebnu vrećicu (spermatoforu), koju ženka odmah hvata rubovima kloake. U ovom slučaju nema seksualnog odnosa, već je oplodnja unutrašnja. Konačno, kod nekih vrsta mužjak ubacuje sjeme u kloaku ženke koristeći svoju izbočenu kloaku.
Kod mnogih vrsta polni dimorfizam je dobro izražen (u boji, građi prednjih nogu kojima mužjaci drže ženke i drugim karakteristikama). Mužjaci brojnih vrsta mogu napraviti vrlo glasni zvuci zahvaljujući pojačavanju ovih zvukova glasovnim vrećicama - rezonatorima.
Razvoj. Razvoj vodozemaca obično se odvija u vodi. Iz oplođenih jaja razvijaju se larve (punoglavci) koje imaju riblji oblik. Dišu kroz škrge i unutrašnja struktura podseća na ribu. Tokom perioda rasta dolazi do transformacije (metamorfoze) punoglavaca: prvo im rastu zadnje noge, zatim atrofiraju prednje noge, škrge i rep (kod bezrepih životinja), razvijaju se pluća, Velike promjene nastaju u cirkulatornom sistemu itd.
Porijeklo. Vodozemci, kao što je gore objašnjeno (str. 296), potječu od riba s režnjevima. Parne peraje drevnih režnjevitih riba, iz kojih su se razvili zglobni udovi kopnenih kralježnjaka, bile su kratke i široke, sadržavale su mnogo sitnih koštanih elemenata, nepovezanih zglobovima, smještenih u nekoliko (najmanje osam) poprečnih redova. Pojasevi na kojima su počivala peraja bili su relativno slabo razvijeni (posebno karlični pojas). Zbog transformacije peraja u kopnene udove došlo je do značajnih promjena u skeletu.
Najprije su smanjeni mnogi koštani elementi: u prvim proksimalnim redovima ostala je samo jedna kost, u prednjoj nozi - rame, u stražnjoj - butna kost; u drugim redovima - dvije kosti, u prednjoj nozi - radijus i ulna, pozadi - tibija i fibula; u sljedeća dva reda ostalo je pet kostiju koje čine karpus na prednjoj nozi i tarsus na stražnjoj nozi; V sljedeći red preostalih pet kostiju uključeno je u metakarpus prednje noge i metatarzalne kosti zadnje noge; preostala tri reda sa po pet kostiju postala su falange prstiju. Smanjenje broja kostiju doprinijelo je povećanju snage nogu.
Drugo, kosti prva dva reda (odnosno ramena i podlaktice, butine i potkolenice) su bile jako produžene, što je bilo veoma važno za povećanje brzine kretanja.
Treće, razvili su se zglobovi između navedenih kostiju, odnosno udovi su postali zglobni, što je najvažniji uslov za njihov rad.
Četvrto, pojasevi udova su ojačani (vidi gore za opis pojaseva). Paralelno ove promjene dogodile su se duboke promjene na nervnom, mišićnom i vaskularni sistemi noge Promjene u drugim organskim sistemima do kojih je došlo tijekom transformacije riba s režnjevim perajima u vodozemce opisane su u opšte karakteristike ovo drugo.
Najstariji vodozemci bili su stegocefali (Sl. 154), koji su bili brojni u periodu karbona, a konačno su izumrli u trijasu. Živjeli su uz obale akumulacija i provodili dosta vremena u vodi. Glava ovih životinja bila je prekrivena ljuskama, što objašnjava njihovo ime (stegocefali - pokrivene glave). Njihov skelet imao je mnogo primitivnih osobina: koštani elementi nogu bili su mali i neznatno različiti po veličini, pršljenovi bikonkavni, pojasevi udova slabo razvijeni, itd. Od njih su nastale moderne grupe vodozemaca.
Mozakžabe, kao i druge vodozemce, odlikuju se sljedećim karakteristikama u odnosu na ribe:
A) progresivni razvoj mozak, izražen u razdvajanju uparenih hemisfera uzdužnom pukotinom i razvojem sive tvari drevnog korteksa (arhipalijum) u krovu mozga;
b) slab razvoj malog mozga;
c) slaba ekspresija pregiba mozga, zbog čega se srednji i srednjim odjeljenjima jasno vidljivo odozgo.
Dijamantski mozak(rombencefalon)
Medulla oblongata (myelencephalon, medulla oblongata), u koju kičmena moždina prolazi kranijalno, razlikuje se od ove druge po većoj širini i odlasku od bočnih površina velikih korijena stražnjih kranijalnih živaca. Na dorzalnoj površini produžene moždine nalazi se jama u obliku dijamanta (fossa rhomboidea), accommoding četvrta moždana komora (ventriculus quartus). Odozgo je prekriveno tankom vaskularna kapica, koji se briše zajedno sa meninge. Ventralna pukotina, nastavak ventralne pukotine kičmene moždine, prolazi duž ventralne površine produžene moždine. Medulla oblongata sadrži dva para konopca (snopova vlakana): donji par, odvojen ventralnom fisurom, su motorni, gornji par su senzorni. Oblongata medulla sadrži centre vilice i sublingvalni aparat, organ sluha, kao i probavni i respiratorni sistem.
Mali mozak nalazi se ispred romboidne jame u obliku visokog poprečnog grebena kao izdanaka njenog prednjeg zida. Mala veličina malog mozga određena je malom i ujednačenom pokretljivošću vodozemaca - u stvari, sastoji se od dva mala dijela, usko povezana sa akustičnim centrima produžene moždine (ovi dijelovi su očuvani kod sisara kao fragmenti malog mozga (flokule)). Tijelo malog mozga - centar koordinacije s drugim dijelovima mozga - vrlo je slabo razvijeno.
Srednji mozak (mesencephalon) gledano sa leđne strane, predstavljena je sa dva tipična optički režnjevi (lobus opticus s. tectum opticus), imaju izgled parnih jajolikih uzvišenja koje formiraju gornje i bočne dijelove srednjeg mozga. Krov optičkih režnjeva formira siva tvar - nekoliko slojeva nervne celije. Tektum kod vodozemaca je najznačajniji dio mozga. Optički režnjevi sadrže šupljine koje su bočne grane cerebralni (Sylvii) akvadukt (aquaeductus cerebri (Sylvii), koji povezuje četvrtu moždanu komoru sa trećom.
Dno srednjeg mozga formiraju debeli snopovi nervnih vlakana - noge veliki mozak(cruri cerebri), povezuje prednji mozak sa produženom moždinom i kičmenom moždinom.
Prednji mozak(prosencephalon) sastoji se od srednjeg i telencephalon, ležeći u seriji.
Diencephalon vidljiv odozgo kao romb, sa oštrim uglovima usmjerenim na strane.
Dijelovi diencefalona leže oko vertikalno smještene široke pukotine treća moždana komora (ventriculus tertius). Lateralno zadebljanje zidova komore - vizuelni kvržici ili thalamus. Kod riba i vodozemaca, talamus je od sekundarnog značaja (kao koordinirajući senzorni i motorički centri). Membranski krov treće moždane komore - epitalamus ili epithalamus - ne sadrži neurone. Sadrži gornji dio medularna žlezda - epifiza (epifiza). Kod vodozemaca epifiza već služi kao žlijezda, ali još nije izgubila karakteristike parijetalnog organa vida. Ispred epifize diencefalon je prekriven membranoznim krovom, koji se oralno okreće prema unutra i prelazi u prednji horoidni pleksus (koroidni tektum treće komore), a zatim u završnu ploču diencefalona. Inferiorno se komora sužava, formirajući se hipofizni lijevak (infundibulum), donja medularna žlezda je kaudoventralno pričvršćena za nju - hipofiza (hipofiza). Ispred, na granici između dna terminala i međudijelova mozga, nalazi se krst optičkih nerava(chiasma nervorum opticorum). Kod vodozemaca, većina vlakana optičkih živaca ne zadržava se u diencephalonu, već ide dalje - do krova srednjeg mozga.
Telencephalon njegova dužina je gotovo jednaka dužini svih ostalih dijelova mozga. Sastoji se od dva dijela: olfaktornog mozga i dvije hemisfere, odvojene jedna od druge sagitalna (u obliku strelice) pukotina (fissura sagittalis).
Hemisfere telencefalona (haemispherium cerebri) zauzimaju zadnje dvije trećine telencefalona i vise preko prednjeg dijela diencefalona, djelomično ga prekrivajući. Unutar hemisfera postoje šupljine - bočne moždane komore (ventriculi lateralis), kaudalno komunicira sa trećom komorom. U sivoj tvari moždanih hemisfera vodozemaca mogu se razlikovati tri područja: dorsomijalno se nalazi stari korteks ili hipokampus (archipallium, s. hippocampus), lateralno - drevna kora(paleopalium) i ventrolateralno - bazalni gangliji, odgovarajući striata (corpora striata) sisari. Strijatum i, u manjoj mjeri, hipokampus su korelativni centri, pri čemu je potonji povezan sa olfaktorna funkcija. Antička kora je isključivo olfaktorni analizator. Na ventralnoj površini hemisfera uočljivi su žljebovi koji odvajaju striatum od drevnog korteksa.
Olfaktorni mozak (rhinencephalon) zauzima prednji dio telencefalona i formira se mirisni režnjevi (lukovice) (lobus olfactorius), međusobno zalemljeni u sredini. Bočno su odvojeni od hemisfera marginalnom fosom. Mirisni režnjevi sa prednje strane sadrže olfaktorne nerve.
10 pari se proteže od žabljeg mozga kranijalnih nerava. Njihovo formiranje, grananje i zona inervacije se suštinski ne razlikuju od onih kod sisara
Mišićni sistem. Razlikuje se od takve ribe uglavnom po većoj razvijenosti mišića udova i većoj diferencijaciji mišića trupa, koji se sastoji od složenog sistema pojedinačnih mišića. Kao rezultat toga, primarna mišićna segmentacija je poremećena, iako u nekim trbušnim i dorzalnim mišićima i dalje ostaje prilično jasna.
Nervni sistem. Mozak vodozemaca razlikuje se od mozga riba uglavnom po većoj razvijenosti prednjeg mozga, potpunom odvajanju njegovih hemisfera i nerazvijenom malom mozgu, koji je samo mali greben nervne tvari koji prekriva prednji dio četvrte komore. Razvoj prednjeg mozga izražava se ne samo u njegovom povećanju i diferencijaciji, već iu činjenici da, pored dna bočnih komora, njihove strane i krov sadrže nervne supstance, odnosno kod vodozemaca se pojavljuje pravi moždani svod - arhipalijum (od modernih riba plućnjaci imaju arhipalijum). Mirisni režnjevi su samo slabo razgraničeni od hemisfera. Diencephalon je samo malo prekriven odozgo susjednim dijelovima. Parietalni organ je pričvršćen za njegov krov, a od dna se proteže dobro izražen lijevak za koji je pričvršćena hipofiza. Iako je srednji mozak značajan dio, on je relativno manji od onog kod riba. Nerazvijenost malog mozga, kao kod plućnjaka, povezana je s jednostavnošću pokreta tijela: vodozemci su uglavnom sjedilačke životinje, ali kod onih koje, poput žaba, mogu napraviti brze pokrete, ograničeni su na skakanje, odnosno vrlo jednostavne pokrete. Iz mozga, kao koštane ribe, nastaje samo 10 pari nerava glave; XII par (hipoglosalni nerv) se proteže izvan lobanje, a XI par (pomoćni nerv) nije uopće razvijen.

. I - vrh; II - dno; III - strana; IV - u uzdužnom presjeku (prema Parkeru):
1 - hemisfere prednjeg mozga, 2 - mirisni režanj, 3 - njušni nerv, 4 - diencefalon, 5 - vizuelni hijazam, 6 — lijevak, 7 - hipofiza, 8 - srednji mozak, 9 - mali mozak, 10 - produžena moždina, 11 - četvrta komora, 12 - kičmena moždina, 13 - treća komora, 14 - Silvijev akvadukt,
III - X - glavni nervi, XII - hipoglosalni nerv

, shema (prema Grguru):
1 - lobanja, 2 - produžena moždina, 3 - slušni nerv, 4 - polukružni kanali, 5 - šupljina srednjeg uha, 6 - Eustahijeva cijev, 7 - ždrijelo, 8 - uho, 9 - bubna opna
U žabi postoji 10 pari pravih kičmenih živaca. U formaciji učestvuju tri prednja para brahijalni pleksus, inervira prednje udove, i četiri zadnja para - u formiranju lumbosakralnog pleksusa, inervira zadnje udove.
Simpatički nervni sistem žabe, kao i kod svih vodozemaca, veoma je dobro razvijen i predstavljen je uglavnom sa dva nervna stabla koja se protežu sa obe strane kičme i formirana su od lanca nervnih ganglija povezanih jedni s drugima konopcima i povezan sa kičmenim nervima.
Još zanimljivih članaka
U ribi mozak u cjelini je mali. Slabo je razvijen prednji dio. Prednji mozak nije podijeljen na hemisfere. Krov mu je tanak, sastoji se samo od epitelnih ćelija i ne sadrži nervno tkivo. Baza prednjeg mozga uključuje striatum, a od njega se protežu mirisni režnjevi. Funkcionalno, prednji mozak je najviši olfaktorni centar.
U diencefalonu, sa kojim su povezane epifiza i hipofiza, nalazi se hipotalamus, koji je centralni organ endokrinog sistema. Srednji mozak riba je najrazvijeniji. Sastoji se od dvije hemisfere i služi kao najviša vizuelni centar. Osim toga, predstavlja najviši integrirajući dio mozga. Zadnji mozak sadrži mali mozak, koji regulira koordinaciju pokreta. Vrlo je dobro razvijen u vezi sa kretanjem riba u trodimenzionalnom prostoru. Oblongata medulla osigurava komunikaciju između viših dijelova mozga i kičmene moždine i sadrži centre disanja i cirkulacije. Mozak ovog tipa, u kojem je najviši centar integracije funkcija srednji mozak, naziva se ihtiopsid.
Kod vodozemaca mozak je takođe ihtiopsid. Međutim, njihov prednji mozak je velik i podijeljen na hemisfere. Njegov krov se sastoji od nervnih ćelija, čiji se procesi nalaze na površini. Poput ribe, srednji mozak dostiže veliku veličinu, što ujedno predstavlja i najviši integrirajući centar i centar vida. Mali mozak je donekle smanjen zbog primitivne prirode pokreta. Uslovi zemaljskog postojanja reptili zahtijevaju složeniju morfofunkcionalnu organizaciju mozga. Prednji mozak je najveći dio u odnosu na ostatak. U njemu je posebno razvijen striatum. Na njih se prenose funkcije najvišeg integrativnog centra. Ostrva kore vrlo primitivne strukture pojavljuju se po prvi put na površini krova drevni - archicortex. Srednji mozak gubi svoju važnost kao vodeći dio, a njegova relativna veličina se smanjuje. Mali mozak je visoko razvijen zbog složenosti i raznovrsnosti pokreta gmizavaca. Mozak ovog tipa, u kojem vodeći dio predstavljaju striatalna tijela prednjeg mozga, naziva se sauropsid.
Kod sisara - sisara tip mozga. Odlikuje se snažnim razvojem prednjeg mozga zbog korteksa, koji se razvija na bazi malog ostrva korteksa reptila i postaje integrativni centar mozga. Sadrži najviše centre vizuelnog, slušnog, taktilnog, motorni analizatori, kao i centri više nervne aktivnosti. Kora ima vrlo složena struktura i zove se nova kora - neokorteks. Sadrži ne samo tijela neurona, već i asocijativna vlakna koja povezuju njegove različite dijelove. Također je karakteristično prisustvo spojnice između obje hemisfere, u kojoj se nalaze vlakna koja ih međusobno povezuju. Diencefalon, kao i druge klase, uključuje hipotalamus, hipofizu i epifizu. U srednjem mozgu nalazi se kvadrigemina u obliku četiri tuberkula. Dva prednja su povezana vizuelni analizator, dva zadnja - sa slušnim.
Glavne faze evolucije centralnog nervni sistem odražavaju se i na ontogenezu čovjeka. U fazi neurulacije, neuralna ploča se polaže, pretvarajući se u žlijeb, a zatim u cijev. Prednji kraj cijevi prvo formira tri moždane vezikule: prednju, srednju i stražnju. . Nakon toga, prednja vezikula se dijeli na dva, diferencirana na prednji mozak i diencefalon - srednja moždana vezikula se razvija u srednji mozak, a stražnja vezikula u zadnji mozak i produženu moždinu.
Monogeni tip nasljeđivanja. Primjeri.
Poligene ili multifaktorske bolesti. Osobine nasljeđivanja.
Vodozemci (vodozemci).
Prednji mozak vodozemci formiraju dvije hemisfere, unutar kojih se nalaze lateralni ventrikuli sa horoidnim pleksusima. Ispred prednjeg mozga leže velike mirisne lukovice. Slabo su odvojeni od hemisfera i kod bezrepih vodozemaca rastu zajedno duž srednje linije. Signali koji dolaze iz olfaktornih lukovica analiziraju se u prednjem mozgu, koji je u suštini najviši olfaktorni centar. Krov prednjeg mozga formira primarni cerebralni svod - arhipalijum. Sadrži nervna vlakna ( bijele tvari), a u dubini, ispod njih, leže nervne ćelije. Na dnu prednjeg mozga nalaze se nakupine neurona - strijatna tijela.
Neposredno iza hemisfera prednjeg mozga nalazi se diencephalon sa dobro razvijenim gornjim moždanim dodatkom - epifizom i donjim moždanim dodatkom - hipofizom. Uopšteno govoreći, diencephalon je sličan onom kod riba.
Najveći dio mozga kod vodozemaca je srednji mozak. Izgleda kao dvije hemisfere prekrivene korom. Uključuje optički trakt kao nastavak optičkih nerava i tu je vizuelni analizator integrisan sa drugim senzorni putevi i formira se centar koji obavlja složene asocijativne funkcije. Dakle, srednji mozak služi kao vodeći odjel centralnog nervnog sistema, gdje se analiziraju primljene informacije i generišu impulsi odgovora, tj. Vodozemci, kao i ribe, imaju ihtiopsidni tip mozga.
Rice. 7. Mozak žabe (dorzalni pogled).
1 – hemisfere prednjeg mozga.
2 – mirisni režnjevi.
3 – olfaktorni nervi.
4 – diencephalon.
5 – srednji mozak.
6 – mali mozak.
7 – produžena moždina.
8 – kičmena moždina.Mali mozak kod većine repatih i bezrepih vodozemaca male veličine i ima izgled poprečnog grebena na prednjem rubu romboidne jame produžene moždine. Slab razvoj malog mozga odražava nesofisticiranu motoričku koordinaciju vodozemaca. Veći dio malog mozga sastoji se od srednjeg dijela (cerebelarnog tijela), gdje su integrirani signali iz mišićnih receptora i vestibularnog sistema.
Kod vodozemaca, kao i kod riba, cerebelarna nervna vlakna su povezana sa srednjim mozgom, moždanim deblom i kičmenom moždinom. Vestibularno-cerebelarne veze određuju sposobnost životinja da koordiniraju pokrete tijela.
Medulla po osnovnim karakteristikama sličan produženoj moždini ribe. Postoji 10 pari kranijalnih nerava koji potiču iz moždanog stabla.
Reptili (gmizavci).
Gmazovi su prave kopnene životinje koje mogu živjeti, razmnožavati se i razvijati daleko od vodenih tijela. Pripadaju višim kičmenjacima. Njihov nervni sistem je, zbog aktivnog i složenog načina života, bolje razvijen od nervnog sistema vodozemaca.

Rice. 8. Mozak guštera (prema Parkeru).
A - pogled odozgo.
B – pogled odozdo.
B – pogled sa strane.
1 – prednji mozak; 2 – strijatum; 3 – srednji mozak; 4 – mali mozak; 5 - produžena moždina; 6 – lijevak; 7 – hipofiza; 8 - hijazma; 9 – mirisni režnjevi; 10 – hipofiza; II – XII- nervi glave.Prednji mozak znatno veće veličine od vodozemaca i ima složeniju strukturu; povećava se njihova sposobnost formiranja uslovnih refleksa, brže se uspostavljaju nove veze sa spoljašnjim okruženjem i bolje se prilagođavaju promenama sredine nego njihovi preci. Prednji mozak se sastoji od dvije hemisfere, koje rastući unazad pokrivaju diencefalon sa izuzetkom epifize i parijetalnog organa. Proširenje prednjeg mozga nastaje uglavnom zbog striatalnih tijela (klastera neurona) smještenih u području dna bočnih ventrikula. Oni djeluju kao viši integrativni centar, obezbjeđujući analizu informacija koje ulaze u prednji mozak i razvoj odgovora. Time prestaje biti samo olfaktorni centar. Ova vrsta mozga se zove sauropsid. Što se tiče moždanog svoda, u njemu se dešavaju važne transformacije. U obje hemisfere krova prednjeg mozga, po prvi put u evoluciji, pojavljuju se dva otoka sive tvari (kortikalni rudimenti) - jedan se nalazi na medijalnoj, a drugi na bočnoj strani hemisfera. Funkcionalno je značajna samo medijalna insula, koja predstavlja najviši olfaktorni centar. Općenito, otoci korteksa imaju primitivnu strukturu i nazivaju se drevni korteks (arhikorteks). Većina autora smatra da su ostrva korteksa jednoslojna, iako se kod krokodila mogu razlikovati dva ili čak tri sloja.
Mirisni režnjevi povezani s prednjim mozgom su dobro razvijeni. Kod nekih vrsta zauzimaju sjedeći položaj, ali se češće diferenciraju na lukovicu i stabljiku.
Istraživanje prednjeg mozga reptila je bitan za evolucijsku neurohistologiju, jer oni su ključna točka u evoluciji kralježnjaka, od kojih je razvoj prednjeg mozga tekao u dva fundamentalno različita smjera: duž striatalnog puta s dominantnim razvojem subkortikalnih struktura kod ptica i duž kortikalnog puta s dominantnim razvojem kortikalnih struktura kod sisara.
Diencephalon na tankom krovu ima dvije mjehuraste formacije, od kojih se jedna nalazi ispred i naziva se parijetalni ili parapinealni organ, a druga iza je epifiza (pinealna žlijezda). Parapinealni organ obavlja fotosenzitivnu funkciju, pa se stoga naziva i parijetalno oko. Zapravo, parapinealni organ i epifiza čine tandem, koji je regulator dnevne aktivnosti životinja. Međutim, parijetalni organ se ne nalazi kod svih gmizavaca. U takvim slučajevima, mehanizam za regulisanje dnevne aktivnosti će biti drugačiji: informacije o dužini dnevnim satima ne dolazi iz parapinealnog organa, već iz vizuelni sistem.
Srednji mozak predstavljen kolikulusom i po svojim glavnim karakteristikama ima istu organizaciju koja je karakteristična za vodozemce, međutim, gmizavce karakteriše precizniji topografski prikaz svakog od senzornih sistema u srednjem mozgu. Štaviše, skoro sve motorna jezgra mali mozak i duguljasta moždina stupaju u interakciju sa neuronima krova srednjeg mozga. Istovremeno, srednji mozak gubi svoj značaj kao glavni integrativni odjel centralnog nervnog sistema. Ova funkcija prelazi na prednji mozak.
Neka od vizuelnih i slušnih vlakana šalju se obilaznim putevima, zaobilazeći srednji mozak, do prednjeg mozga. U srednjem mozgu, centri za osiguranje automatskih urođenih reakcija tijela, dobiveni su čak ranim fazama evolucija kičmenjaka. Novi centri prednjeg mozga preuzimaju funkcije tekuma i formiraju nove motoričke puteve.
Mali mozak zbog razvoja hodanja i trčanja kod gmizavaca, bolje je razvijen nego kod vodozemaca. Sastoji se od centralnog aksijalnog dijela, nazvanog crv, a neki imaju bočne režnjeve. Mali mozak karakteriziraju brojne veze s drugim dijelovima nervnog sistema koji se odnose na lokomociju. Ovdje povezan spinocerebelarni trakt, prisutan i kod riba i kod vodozemaca, proteže se do dorzalnog dijela vermisa. Postoje cerebelarne veze sa vestibularnim aparatom, srednjim mozgom i rombencefalonom. Vestibularno-cerebelarne veze kontrolišu položaj tijela u prostoru, a talamičke veze regulišu tonus mišića.
Medulla formira oštar zavoj u okomitoj ravni, karakterističan općenito za amniote.
Postoji 12 pari kranijalnih nerava koji potiču iz moždanog stabla.
Ptice.
Ptice su visoko specijalizirana grupa kralježnjaka prilagođena letu. Budući da su fiziološki bliske gmizavcima, ptice imaju mozak na mnogo načina sličan mozgu gmizavaca, iako se razlikuju po nizu progresivnih karakteristika, što je dovelo do složenijeg i raznovrsnijeg ponašanja. Mozak im je prilično velik. Ako je kod gmizavaca njegova masa približno jednaka masi kičmene moždine, onda je kod ptica uvijek veća. Pregibi mozga su oštro izraženi.
Povećanje mozga prvenstveno je posljedica razvoja prednji mozak, koji u suštini pokriva sve dijelove mozga s izuzetkom malog mozga. Zbog toga se srednji mozak ne vidi odozgo, iako je dobro razvijen. Krov prednjeg mozga ostaje slabo razvijen. Cora ne samo da ne prima dalji razvoj, ali u hemisferama lateralno ostrvo korteksa nestaje, a medijalno zadržava značaj višeg olfaktornog centra.
Povećanje veličine prednjeg mozga nastaje zbog dna, gdje se nalaze velika striatalna tijela, koja su vodeći dio mozga. To jest, ptice zadržavaju sauropsid tip mozga.
Mirisni režnjevi su vrlo mali, što je povezano sa slabim razvojem čula mirisa, te su usko uz prednji mozak.
Diencephalon mala. Kod većine ptica obično je očuvana samo epifiza, a parapinealni organ nestaje u kasnom embrionalnom periodu. Informacije o dužini dnevnog svetla ne dolaze iz parapinealnog organa, već direktno iz vizuelnog sistema. U talamusu je najrazvijeniji dorzalni dio, koji je preklopni centar aferentnih veza s prednjim mozgom. Sadrži kompleks jezgara na putu silaznih vlakana iz motoričkih centara prednjeg mozga. Ispod njega je hipotalamus, povezan sa hipofizom, koja je kod ptica dobro razvijena. Hipotalamus igra glavnu ulogu u hormonskoj regulaciji organizma, održavanju homeostaze, seksualnog ponašanja i ponašanja u ishrani.
Srednji mozak sastoji se od dvije velike humke. Sadrži najviše centre za obradu vizuelnih i slušnih informacija, kao i centre za regulisanje vrsta specifičnih naslednih oblika koordinisanih reakcija, koji su osnova života. Postoje i senzorna jezgra koja obavljaju asocijativnu funkciju, šaljući signale u diencefalon i prednji mozak. Istovremeno, dolazi do smanjenja broja silaznih veza između krova srednjeg mozga i motornih centara zbog pojave kod ptica striatalno-retikularnih relativno autonomnih veza između striatalnih tijela prednjeg mozga i retikularna formacija moždano stablo.
Mali mozak velika i njena struktura postaje složenija. Sprijeda dolazi u dodir sa stražnjim rubovima hemisfera prednjeg mozga, a iza pokriva značajan dio produžene moždine. Mali mozak je podijeljen na srednji dio (vermis) i bočne projekcije. Njegov je crv prošaran karakterističnim poprečnim žljebovima. Relativno složena struktura malog mozga nastaje zbog složenih pokreta koji zahtijevaju visoku koordinaciju tokom leta.Medulla relativno mala, njegova donja strana, čak i više nego kod gmizavaca, čini zavoj prema dolje, au području diencephalona nalazi se zavoj prema gore.
Kranijalni nervi ptica predstavljeni su u 12 parova.
sisari.
Sisavci su najorganizovanija klasa kičmenjaka sa visoko razvijenim centralnim nervnim sistemom. U tom smislu, adaptivne reakcije sisara na uslove okoline su složene i veoma napredne.
Prednji mozak velika, značajno je veća od svih ostalih dijelova mozga. Njegove hemisfere rastu u svim smjerovima, skrivajući diencephalon. Srednji mozak je izvana vidljiv samo kod aplacentala i donjih placenti, a kod kopitara, mesoždera, kitova i primata prekriven je stražnjim dijelom moždane hemisfere. Kod antropoida i ljudi okcipitalni režnjevi prednji mozak se gura na mali mozak.
Ako se u početku, tokom evolucije, glavnina telencefalona sastojala od njušnih režnjeva, onda su kod sisara samo niži razvili mirisne režnjeve, a u višim njušni režnjevi imaju oblik malih dodataka, podijeljenih na njušne režnjeve i olfaktornog trakta.
Povećanje relativne veličine prednjeg mozga sisara povezano je prvenstveno s rastom njegovog krova, a ne striatuma, kao kod ptica. Medularni svod (krov) je formiran od sive tvari koja se zove korteks. Potonji je kompleks koji se sastoji od antičkog ogrtača (paleopalium), starog ogrtača (archipallium) i novog ogrtača (neopalium). Novi ogrtač zauzima srednji položaj, smješten između starog i antičkog ogrtača. Stari plašt, ili stari korteks, nalazi se medijalno i u prošlosti se zvao hipokampus ili Amonov rog. Drevni ogrtač, ili drevna kora, zauzima bočni položaj.
Novi ogrtač se obično naziva neokorteks ( neokorteks) i od toga se uglavnom sastoje hemisfere prednjeg mozga. U ovom slučaju, površina hemisfera može biti glatka (lisencefalična) ili presavijena (sa žljebovima i zavojima). Osim toga, bez obzira na to, u hemisferama se razlikuje od 4 do 5 režnjeva. Princip podjele prednjeg mozga na režnjeve temelji se na topografiji određenih žljebova i konvolucija. Podjela na režnjeve u lisencefalnom (glatkom) mozgu je uslovna. Obično postoje parijetalni režnjevi, temporalni, okcipitalni i frontalni, a kod viših primata i ljudi postoji i peti režanj koji se naziva insula. Nastaje u embrionalnom periodu zbog rasta temporalnog režnja na ventralnu stranu hemisfera.Uzimajući lisencefalni mozak kao početni tip moždanih hemisfera, razlikuju se tri opcije za razvoj obrasca žljebova: uzdužni, lučni i "tip primata". Kod tipa primata žljeb u frontalnim režnjevima je usmjeren rostralno, a u temporalnim režnjevima - ventro-dorzalno
Na položaj brazdi i vijuga može značajno uticati oblik mozga. Kod većine sisara mozak je izdužen u rostro-kaudalnom smjeru. Međutim, kod mnogih dupina mozak je bočno proširen i relativno skraćen.
Za karakterizaciju prednjeg mozga sisara veliki značaj, pored brazdi i konvolucija, ima obrazac distribucije u neuronskom korteksu (citoarhitektura). Neokorteks sisara ima šestoslojnu strukturu i karakteriše ga prisustvo piramidalnih ćelija, kojih nema u mozgu drugih kralježnjaka. Posebno velike piramidalne ćelije (Betz ćelije) nalaze se u motornom korteksu. Njihovi aksoni prenose nervne impulse do motornih neurona kičmene moždine i motornih neurona motornih jezgara kranijalnih nerava.
Različita područja moždane kore su specijalizirana područja za obradu informacija koje dolaze iz različitih osjetilnih organa. Postoje senzorna i motorička područja. Potonji formiraju silazne puteve nervnih vlakana do moždanog stabla i motornih jezgara kralježnice. Između osjetljivih i motoričkih područja korteksa postoje integrativna područja koja kombinuju ulaze senzornih i motoričkih područja korteksa i određuju izvođenje specijaliziranih funkcija specifičnih za vrstu. Osim toga, postoje asocijativne zone korteksa koje nisu povezane sa specifičnim analizatorima. Oni predstavljaju nadgradnju nad preostalim područjima korteksa, obezbeđujući misaone procese i skladištenje specifične i individualne memorije.
Čitav kompleks zona raspoređenih u korteksu povezan je sa funkcionalnom specijalizacijom polja. U ovom slučaju, morfološke i funkcionalne granice polja prilično se poklapaju. Kriterij za identifikaciju određenog polja je promjena u distribuciji ćelijskih elemenata u korteksu ili pojava novog podsloja u njemu.
Odlike arhitektonike pojedinih oblasti morfološki su izraz njihove funkcionalne specijalizacije. Razlog za promjenu citoarhitektonike u poljima je povećanje broja uzlaznih i silaznih nervnih vlakana. Sada su stvorene topološke karte polja za ljude i za mnoge laboratorijske životinje.
Polja kore velikog mozga su dio određenih režnjeva i sama su podijeljena u funkcionalne zone povezane s određenim organima ili njihovim dijelovima i imaju uređenu unutrašnju strukturu. U svakom polju ili zoni postoje tzv moduli vertikalna urednost organizacije korteksa. Modul ima ili oblik stupca ili glomerulusa, koji uključuje neurone smještene po cijeloj debljini korteksa. Kolona sadrži grupu od 110 neurona koji se nalaze između para kapilara koji se protežu preko prečnika korteksa.
U fazi formiranja mozga najstarijih hominida, područje na koje je bilo usmjereno djelovanje prirodne selekcije bio je korteks i, prije svega, njegovi sljedeći dijelovi: donja parijetalna, donja frontalna i temporo-parijetalna regija. Prednost preživljavanja otišla je tim pojedincima, a potom i onim populacijama ljudi u nastajanju za koje se pokazalo da su napredni u smislu razvoja nekih elemenata dijelova korteksa ( veća površina polja sa raznovrsnijim i mobilnim vezama, poboljšanim uslovima cirkulacije, itd.). razvoj novih veza i struktura u korteksu pružio je nove mogućnosti za proizvodnju alata i team building. Zauzvrat, novi nivo tehnologije, počeci kulture i umjetnosti kroz prirodnu selekciju doprinijeli su razvoju mozga.
Do danas je formirana ideja o specifičnom sistemskom kompleksu korteksa ljudskog prednjeg mozga, uključujući inferiorni parijetalni, stražnji gornji temporalni i donji frontalni režnjevi korteksa. Ovaj kompleks je povezan sa višim funkcijama - govorom, radnom aktivnošću i apstraktnim mišljenjem. Generalno, to je morfološki supstrat drugog signalnog sistema. Ovaj sistem nema svoje periferne receptore, već koristi stari receptorski aparat raznih čulnih organa. Na primjer, utvrđeno je da jezik ima poseban dio taktilnog aparata, čiji razvoj određuje slijed proizvodnje zvuka u početnim fazama formiranja djetetovog artikuliranog govora.
Submantilne strukture prednjeg mozga uključuju bazalne ganglije, striatum (drevni, stari i novi) i septalno polje.
U različitim dijelovima prednjeg mozga i diencefalona nalazi se kompleks morfofunkcionalnih struktura tzv limbički sistem. Potonji ima brojne veze sa neokorteksom i autonomnim nervnim sistemom. Integrira moždane funkcije kao što su emocije i pamćenje. Uklanjanje dijela limbičkog sistema dovodi do emocionalne pasivnosti životinje, a njegova stimulacija dovodi do hiperaktivnosti. Najvažnija funkcija Limbički sistem je u interakciji sa memorijskim mehanizmima. Kratkoročno pamćenje je povezano sa hipokampusom, a dugotrajno sa neokorteksom. Kroz limbički sistem, individualno iskustvo životinje se izvlači iz neokorteksa, dolazi do motoričke kontrole unutrašnjih organa i hormonske stimulacije životinje. Štoviše, što je niži stupanj razvoja neokorteksa, ponašanje životinje više ovisi o limbičkom sistemu, što dovodi do dominacije emocionalno-hormonske kontrole nad donošenjem odluka.
Kod sisara, silazne veze od neokorteksa do limbičkog sistema omogućavaju integraciju širokog spektra senzornih inputa.
Pojavom prvih rudimenata korteksa kod gmizavaca, mali snop nervnih vlakana koji povezuju lijevu i desnu hemisferu odvojio se od komisure plašta. Kod placentnih sisara takav je snop vlakana mnogo razvijeniji i naziva se corpus callosum. Potonji pruža funkciju međuhemisfernih komunikacija.
Diencephalon, kao i drugi kralježnjaci, sastoji se od epitalamusa, talamusa i hipotalamusa.
Razvoj neokorteksa kod sisara doveo je do naglog povećanja talamusa, a prije svega dorzalnog. Talamus sadrži oko 40 jezgara, u kojima se uzlazni putevi prebacuju na posljednje neurone, čiji aksoni dopiru do kore velikog mozga, gdje se obrađuju informacije koje dolaze iz svih senzornih sistema. Istovremeno, prednja i lateralna jezgra obrađuju i prenose vizuelne, slušne, taktilne, gustatorne i interoceptivne signale do odgovarajućih projekcijske zone kora. Postoji mišljenje da se osjetljivost na bol ne projektuje u korteks hemisfera prednjeg mozga, već centralni mehanizmi nalaze se u talamusu. Ova pretpostavka se zasniva na činjenici da iritacija različitih područja korteksa ne uzrokuje bol, dok iritacija talamusa uzrokuje bol. jak bol. Neki od jezgara talamusa se mijenjaju, a drugi dio su asocijativni (od njih postoje putevi do asocijativnih zona korteksa). U medijalnom dijelu talamusa nalaze se jezgre koje uz niskofrekventnu električnu stimulaciju izazivaju razvoj inhibitornih procesa u moždanoj kori, što dovodi do spavanja. Visokofrekventna stimulacija ovih jezgara uzrokuje djelomičnu aktivaciju kortikalnih mehanizama. Dakle, talamokortikalni regulatorni sistem, kontrolišući protok uzlaznih impulsa, učestvuje u organizovanju promene između sna i budnosti.
Ako se kod nižih kralježnjaka viši senzorni i asocijacijski centri nalaze u srednjem mozgu, a dorzalni talamus je skroman integrator između srednjeg mozga i olfaktorni sistem, tada je kod sisara najvažniji centar za prebacivanje slušnih i somatosenzornih signala. Istovremeno, somatosenzorno područje postalo je najistaknutija formacija diencefalona i igra veliku ulogu u koordinaciji pokreta.
Treba napomenuti da se kompleks talamičkih jezgara formira kako zbog primordija diencefalona, tako i zbog migracije iz srednjeg mozga.
Hipotalamus formira razvijene bočne izbočine i šuplju stabljiku - lijevak. Potonji se završava u stražnjem smjeru s neurohipofizom čvrsto povezanom s adenohipofizom.
Hipotalamus je najviši regulatorni centar endokrine funkcije tijelo. Kombinira endokrine regulatorne mehanizme sa nervnim. Osim toga, to je najviši centar simpatičkog i parasimpatičkog odjela autonomnog nervnog sistema.
Epitalamus služi kao neurohumoralni regulator dnevne i sezonske aktivnosti, koji se kombinuje sa kontrolom puberteta kod životinja.
Srednji mozak formira kvadrigeminalnu regiju, čiji su prednji tuberkuli povezani sa vizuelnim analizatorom, a zadnji sa slušnim. Po omjeru relativnih veličina prednjih i stražnjih tuberkula može se prosuditi koji od sistema, slušni ili vidni, prevladava. Ako su prednji tuberkuli bolje razvijeni, to znači vizualna aferentacija (papkari, mnogi grabežljivci i primati), ako stražnji, onda slušna aferentacija (delfini, slepi miševi itd.).
Tagment je podijeljen na senzornu i motoričku zonu. Motorna zona sadrži motorna jezgra kranijalnih nerava i silazna i uzlazna spinocerebralna vlakna.
U vezi s razvojem neokorteksa kod sisara kao višeg integrativnog centra, urođene reakcije srednjeg mozga omogućile su korteksu da se "ne zahvaća" primitivne forme vrste specifične reakcije na vanjske signale, dok složene asocijativne funkcije preuzimaju specijalizirana kortikalna polja.
Mali mozak kod sisara poprima najsloženiju strukturu. Anatomski se može podijeliti na srednji dio - vermis, hemisfere koje se nalaze s obje njegove strane i flokulonodularne režnjeve. Potonji predstavljaju filogenetski drevni dio - Archicerebellum. Hemisfere su zauzvrat podijeljene na prednje i stražnje režnjeve. Prednji režnjevi hemisfera i stražnji dio cerebelarnog vermisa predstavljaju filogenetski stari mali mozak - paleocerebellum. Filogenetski, najmlađi dio malog mozga, neocerebellum, uključuje prednji dio stražnjih režnjeva hemisfera malog mozga.
Hemisfere malog mozga dijele se na gornju površinu, koja formira koru malog mozga, i nakupine nervnih ćelija - jezgra malog mozga. Kora malog mozga izgrađena je po jednom principu i sastoji se od 3 sloja. Mali mozak je povezan sa ostalim dijelovima centralnog nervnog sistema pomoću tri para pedunula formiranih od snopova nervnih vlakana. Zadnje noge se prvenstveno sastoje od proprioceptivnih vlakana koja dolaze iz kičmene moždine. Srednji pedunci se sastoje od vlakana koja povezuju mali mozak i korteks prednjeg mozga, a prednji pedunci su formirani silažnim vlaknima koja povezuju mali i srednji mozak.Vestibulocerebelarne veze određuju sposobnost životinja da koordiniraju pokrete tijela, što je glavna funkcija arhicerebeluma. Osim toga, novi, snažniji cerebelarni putevi su se formirali kod sisara zbog pojave zupčastog jezgra malog mozga. Prima vlakna iz različitih dijelova hemisfere malog mozga i prenosi signale u talamus, gdje se senzomotorički signali integriraju s aktivnošću kortikalnih centara prednjeg mozga
... funkcionalan filogenija morpho-funkcionalan... centralno nervozan, endokrini, respiratorni i drugi sistemaFunkcionalni država...
Od fundamentalnih nauka morfologije, fiziologije, fizike, evolucijske nastave - do ekologije inteligentnih sistema, medicine, javnog zdravlja (1)
Book... funkcionalan, parametarske, sinergijske karakteristike filogenija i ontogeneza ljudskog mozga. Zbirka znanja o morpho-funkcionalan... centralno nervozan, endokrini, respiratorni i drugi sistemaFunkcionalni država...
Od fundamentalnih nauka morfologije, fiziologije, fizike, evolucijske nastave - do ekologije inteligentnih sistema, medicine, javnog zdravlja (2)
Book... funkcionalan, parametarske, sinergijske karakteristike filogenija i ontogeneza ljudskog mozga. Zbirka znanja o morpho-funkcionalan... centralno nervozan, endokrini, respiratorni i drugi sistemaFunkcionalni država...
Fiziologija centralnog nervnog sistema (1)
Dokumentkodiranje - transformacija informacije u... beskičmenjaci i niže kralježnjaciživotinje... vezu nervozan I endokrinisistema. ... funkcionalan sistema koji primaju signale od organiosjecanja nakon prethodne obrade u central nervozan ...
Boris Gurjevič Meščerjakov Vladimir Petrovič Zinčenko Veliki psihološki rečnik Sadržaj
Dokument... organiosjecanja(I ljudsko tijelo općenito) u kombinaciji sa funkcionalan asimetrija... nervozanćelije ( nervozan G.) izvan c. n. With. U kralježnjaciživotinje G. nalaze se duž kičmene moždine i u zidovima unutrašnjeg organi. U beskičmenjaci ...


